Ostrovní kůra - Insular cortex
| Ostrovní kůra | |
|---|---|
 Izolace pravé strany, odkrytá odstranění opercula. | |
 Koronální část mozku okamžitě dovnitř před mostem. (Insula labeled at right right.) | |
| Detaily | |
| Část | mozková kůra z mozek |
| Tepna | Střední mozek |
| Identifikátory | |
| latinský | Cortex insularis |
| NeuroNames | 111 |
| NeuroLex ID | birnlex_1117 |
| TA98 | A14.1.09.149 A12.2.07.053 |
| TA2 | 5502 |
| FMA | 67329 |
| Anatomické pojmy neuroanatomie | |
The ostrovní kůra (taky ostrov a ostrovní lalok) je část mozková kůra složené hluboko uvnitř laterální sulcus (trhlina oddělující temporální lalok z temenní a čelní laloky ) v každém z nich polokoule z savčí mozek.
Předpokládá se, že jsou součástí insulae vědomí a hrát roli v různých funkcích, s nimiž obvykle souvisí emoce nebo regulace těla homeostáza. Mezi tyto funkce patří soucit a empatie, chuť, vnímání, ovládání motoru, sebeuvědomění, kognitivní fungování, a mezilidská zkušenost. V souvislosti s nimi je zapojen do psychopatologie.
Ostrovní kůra je rozdělena na dvě části: přední insula a zadní insula, ve kterých bylo identifikováno více než tucet polních oblastí. Kortikální oblast překrývající izolaci směrem k bočnímu povrchu mozku je operculum (význam víčko). Opercula jsou tvořeny z částí obklopujících čelních, časových a temenních laloků.
Struktura
Připojení
Přední část ostrova je rozdělena mělkými sulci na tři nebo čtyři krátký gyri.
Přední insula přijímá přímý výstupek z bazální části ventrální mediální jádro thalamu a obzvláště velký příspěvek z centrální jádro amygdaly. Kromě toho přední ostrov sám promítá do amygdala.
Jedna studie na opicích rhesus odhalila rozsáhlá vzájemná spojení mezi ostrovní kůrou a téměř všemi podjádry amygdaloidního komplexu. Zadní insula vyčnívá převážně do dorzálního aspektu laterálního a do centrálních amygdaloidních jader. Naproti tomu přední insula vyčnívá do přední amygdaloidní oblasti a také do mediálního, kortikálního, pomocného bazálního magnocelulárního, mediálního bazálního a laterálního amygdaloidního jádra.[1]
Zadní část ostrova je tvořena a dlouhý gyrus.
Zadní ostrov se vzájemně spojuje s sekundární somatosenzorická kůra a přijímá vstup z spinotalamicky aktivován ventrální posterior inferior thalamová jádra. Ukázalo se také, že tato oblast přijímá vstupy z ventromediálního jádra (zadní části) thalamu, které jsou vysoce specializované na zprostředkování homeostatických informací, jako je bolest, teplota, svědění, místní stav kyslíku a smyslný dotek.[2]
Studie lidského neuroimagingu s využitím difúzní tenzorové zobrazování odhalilo, že přední insula je propojena s oblastmi v temporálním a okcipitálním laloku, operulární a orbitofrontální kůře, trojúhelníkové a operkulární části dolního frontálního gyrusu. Stejná studie odhalila rozdíly v anatomických vzorcích spojení mezi levou a pravou hemisférou.[3]
'kruhový sulcus of insula ' (nebo sulcus z Reilu[4]) je půlkruhový sulcus nebo trhlina[4] která odděluje izolát od sousedních gyri z operculum[5] vpředu, nahoře a za nimi.[4]
Cytoarchitektura
Ostrovní kůra má oblasti proměnlivé buněčné struktury nebo cytoarchitektura, přecházející z zrnitý v zadní část na agranulární v přední část. Ostrov také dostává diferenciál kortikální a thalamský vstup po jeho délce. Přední ostrovní kůra obsahuje populaci vřetenové neurony (také zvaný von Economo neurony), identifikovaný jako charakterizující výrazný podoblast jako agranulární čelní ostrov.[6]
Rozvoj
Ostrovní kůra je považována za samostatnou lalok z telencephalon některými orgány.[7] Jiné zdroje považují ostrov za součást temporální lalok.[8] To je také někdy seskupeno s limbickými strukturami hluboko v mozku do a limbický lalok.[Citace je zapotřebí ] Jako paralimbická kůra je ostrovní kůra považována za relativně starou strukturu.
Funkce
Multimodální senzorické zpracování, senzorická vazba
Studie funkčního zobrazování ukazují aktivaci ostrova během audiovizuálních integračních úkolů.[9][10]
Chuť
Přední ostrov je součástí primární chuťová kůra.[11][12]
Interoceptivní povědomí
Existují důkazy, že kromě základních funkcí může insula hrát roli v některých funkcích vyšší úrovně, které fungují pouze u lidí a jiných lidoopi. Vřetenové neurony nalezené při vyšší hustotě v pravé frontální ostrovní kůře se také nacházejí v přední cingulární kůra, což je další region, který dosáhl vysoké úrovně specializace na lidoopy. Spekulovalo se, že tyto neurony jsou součástí poznávací -emocionální procesy, které jsou specifické pro primáty, včetně lidoopů, jako je empatie a metakognitivní emocionální pocity. To je podporováno výsledky funkčního zobrazování, které ukazují, že struktura a funkce pravé čelní izolace souvisí se schopností cítit vlastní tlukot srdce nebo vcítit se do bolesti druhých. Předpokládá se, že tyto funkce se neliší od funkcí na nižší úrovni ostrova, ale spíše vznikají jako důsledek role ostrova v přenosu homeostatických informací do vědomí.[13][14] Pravá přední izolace pomáhá interoceptivní povědomí o stavech těla, jako je schopnost měřit vlastní čas tlukot srdce. Navíc větší pravý přední ostrovní šedá hmota objem koreluje se zvýšenou přesností v tomto subjektivním smyslu pro vnitřní tělo a s negativním emocionálním zážitkem.[15] Podílí se také na kontrole krevní tlak,[16] zejména během a po cvičení,[16] a její aktivita se liší podle množství úsilí, které člověk věří, že vyvíjí.[17][18]
Ostrovní kůra je také místem, kde je pocit bolest je posuzován podle jeho stupně.[19] Dále je to ostrov, kde si člověk představuje bolest při pohledu na obrazy bolestivých událostí, zatímco přemýšlí o tom, že se stalo s vlastním tělem.[20] Ti s syndrom dráždivého tračníku mít neobvyklé zpracování viscerální bolest v ostrovní kůře související s dysfunkční inhibicí bolesti v mozku.[21]
Další vnímání pravé přední insula je míra nebolestivé teplo[22] nebo bezbolestný chlad[23] kožního pocitu. Mezi další vnitřní pocity zpracovávané izolací patří žaludek nebo břišní distenze.[24][25] Plný měchýř také aktivuje ostrovní kůru.[26]
Jedna studie zobrazování mozku naznačuje, že nepříjemnost je subjektivně vnímána dušnost je zpracována v pravé lidské přední insula a amygdala.[27]
Zpracování mozkové kůry vestibulární pocity zasahují do ostrova,[28] s malými lézemi v přední ostrovní kůře, které mohou způsobit ztrátu Zůstatek a závrať.[29]
Mezi další neinterceptivní vnímání patří pasivní poslech hudby,[30] smích a pláč[31] empatie a soucit,[32] a jazyk.[33]
Ovládání motoru
Při řízení motoru přispívá k motorickému pohybu rukou a očí,[34][35] polykání,[36] motilita žaludku,[37] a artikulace řeči.[38][39] Bylo identifikováno jako „centrální velitelské“ centrum, které to zajišťuje Tepová frekvence a krevní tlak zvýšení na začátku roku 2006 cvičení.[40] Výzkum na základě konverzace ji spojuje s kapacitou pro dlouhé a složité mluvené věty.[41] Podílí se také na motorickém učení[42] a bylo zjištěno, že hraje roli při zotavení motoru po mrtvici.[43]
Homeostáza
Hraje roli v různých homeostatických funkcích souvisejících se základními potřebami přežití, jako je chuť, viscerální vjem a autonomní kontrola. Ostrov ovládá autonomní funkce prostřednictvím regulace sympatických a parasympatických systémů.[44][45] Hraje roli v regulaci imunitního systému.[46][47][48]
Já
Bylo zjištěno, že hraje roli ve zkušenosti tělesného sebeuvědomění,[49][50] smysl pro agenturu,[51] a pocit vlastnictví těla.[52]
Sociální emoce
Přední ostrov zpracovává smysl člověka hnus oba cítit[53] a na pohled na kontaminaci a zmrzačení[54] - i když si jen představujete zážitek.[55] To je spojeno s a zrcadlový neuron podobné propojení mezi vnějšími a vnitřními zkušenostmi.
V sociální praxi se podílí na zpracování porušování norem,[56] emoční zpracování,[57] empatie,[58] a orgasmy.[59]
Ostrov je aktivní při společenském rozhodování. Tiziana Quarto a kol. měřeno emoční inteligence (EI) (schopnost identifikovat, regulovat a zpracovávat emoce sebe i ostatních) šedesáti tří zdravých subjektů. Použitím fMRI EI byla měřena v korelaci s levou ostrovní aktivitou. Subjektům byly zobrazeny různé obrázky výrazy obličeje a za úkol rozhodnout přiblížit se osobě na obrázku nebo se jí vyhnout. Výsledky úkolu sociálního rozhodování přinesly, že jednotlivci s vysokým skóre EI opustili ostrovní aktivaci při zpracování ustráchaných tváří. Jednotlivci s nízkým skóre EI opustili ostrovní aktivaci při zpracování rozzlobených tváří.[60]
Emoce
Ostrovní kůra, zejména její nejpřednější část, je považována za limbický související kůra. Ostrov se stále více stává středem pozornosti pro svou roli v reprezentaci těla a subjektivním emocionálním zážitku. Zejména, Antonio Damasio navrhl, aby tato oblast hrála roli při mapování viscerálních stavů, které jsou spojeny s emocionálním zážitkem, což vede k pocitům vědomí. Toto je v podstatě neurobiologická formulace myšlenek William James, který jako první navrhl, aby subjektivní emoční zkušenost (tj. pocity) vycházela z interpretace tělesných stavů našeho mozku vyvolaných emočními událostmi. Toto je příklad ztělesněné poznání.
Z hlediska funkce se předpokládá, že ostrov zpracovává konvergentní informace za účelem vytvoření emocionálně relevantní kontext pro smyslový zážitek. Přesněji řečeno, přední insula souvisí spíše s čichová, chuťová, viscero-autonomní a limbická funkce, zatímco zadní ostrov souvisí spíše s sluchově-somesteticko-skeletomotorický funkce. Funkční zobrazovací experimenty odhalili, že ostrov má důležitou roli v bolest zkušenosti a zkušenosti z řady základních emoce, počítaje v to hněv, strach, hnus, štěstí, a smutek.[61]
Přední ostrovní kůra (AIC) je považována za zodpovědnou za emoční pocity, včetně mateřské a romantické lásky, hněvu, strachu, smutku, štěstí, sexuálního vzrušení, znechucení, averze, nespravedlnosti, nerovnosti, rozhořčení, nejistoty,[62] nedůvěra, sociální vyloučení, důvěra, empatie, sochařská krása, „stav sjednocení s Bohem“ a halucinogenní stavy.[63]
Funkční zobrazovací studie také zapojily ostrov do vědomých tužeb, jako je touha po jídle a touha po drogách. Společné pro všechny tyto emocionální stavy je, že každý nějakým způsobem mění tělo a je spojen s vysoce výraznými subjektivními vlastnostmi. Ostrov má dobrou polohu pro integraci informací týkajících se tělesných stavů do kognitivních a emocionálních procesů vyššího řádu. Ostrov přijímá informace z „homeostatických aferentních“ senzorických drah prostřednictvím thalamu a odesílá výstup do řady dalších struktur souvisejících s limbikou, jako je například amygdala, ventrální striatum a orbitofrontální kůra, stejně jako motorické kůry.[64]
Studie s využitím magnetická rezonance zjistil, že pravá přední izolace je u lidí výrazně silnější přemýšlet.[65] Další výzkum mozková aktivita a meditace prokázal nárůst šedé hmoty v oblastech mozku včetně ostrovní kůry.[66]
Další studie využívající morfometrii založenou na voxelech a MRI na zkušených Vipassanové meditující bylo provedeno za účelem rozšíření poznatků Lazara a kol., kteří u zkušených meditujících zjistili zvýšené koncentrace šedé hmoty v této a dalších oblastech mozku.[67]
Nejsilnější důkazy proti kauzální roli mozkové kůry insula v emocích pocházejí od Damasia et al. (2012) [68] který ukázal, že pacient, který utrpěl bilaterální léze mozkové kůry insula, vyjádřil plný počet lidských emocí a byl plně schopen emočního učení.
Nápadnost
Funkční neuroimaging výzkum naznačuje, že ostrov je zapojen do dvou typů nápadnost. Interoceptivní zpracování informací, které spojuje interocepci s emocionální význačností a vytváří subjektivní reprezentaci těla. Jedná se nejprve o přední ostrovní kůru s pregenuální přední cingulární kůra (Brodmann oblast 33 ) a přední a zadní střední cingulární kůry a za druhé generál hlavní síť zabývá se monitorováním prostředí, výběrem odezvy a orientací skeletomotorického těla, která zahrnuje celou ostrovní kůru a kůru ve střední cingulaci.[69]
Alternativním nebo možná doplňkovým návrhem je, že pravý přední ostrov reguluje interakci mezi význačností selektivní pozornost vytvořen k dosažení úkolu (systém hřbetní pozornosti) a význačnosti vzrušení vytvořen, aby se soustředil na příslušnou část prostředí (systém ventrální pozornosti).[70] Tato regulace výběžku může být obzvláště důležitá při náročných úkolech, kde je možná pozornost únava a tak způsobit neopatrné chyby, ale je-li příliš mnoho vzrušení, riskuje, že se tím změní na špatný výkon úzkost.[70]
Sluchové vnímání
Nedávný výzkum naznačuje, že je zapojena ostrovní kůra sluchové vnímání. Odpovědi na zvukové podněty byly získány pomocí intrakraniální EEG záznamy získané od pacientů s epilepsií. Zadní část izolátu vykazovala sluchové odpovědi podobné těm, které byly pozorovány v Heschlův gyrus zatímco přední část reagovala na emoční obsah sluchových podnětů.[71]
Přímé záznamy ze zadní části ostrova ukazovaly reakce na neočekávané zvuky v pravidelných zvukových tocích, což je proces známý jako detekce sluchové odchylky. Vědci pozorovali a nesoulad negativity (MMN) potenciál, dobře známý potenciál související s událostí, stejně jako vysokofrekvenční signály aktivity pocházející z místních neuronů.[72]
Jednoduché sluchové iluze a halucinace byly vyvolány elektrickým funkčním mapováním.[73][71]
Klinický význam
Progresivní expresivní afázie
Progresivní expresivní afázie je zhoršení normálu funkce jazyka což způsobí, že jednotlivci ztratí schopnost plynně komunikovat a zároveň budou schopni porozumět jednotlivým slovům a neporušit jiné mimojazykové poznání. Nachází se v různých degenerativních neurologických stavech včetně Pickova nemoc, onemocnění motorických neuronů, kortikobazální degenerace, frontotemporální demence, a Alzheimerova choroba. Je spojen s hypometabolismem[74] a atrofie levé přední ostrovní kůry.[75]
Závislost
Řada funkčních zobrazovacích studií mozku ukázala, že ostrovní kůra se aktivuje, když jsou uživatelé drog vystaveni vlivům prostředí, které vyvolávají chutě. To bylo prokázáno pro různé léky, včetně kokain, alkohol, opiáty, a nikotin. Navzdory těmto zjištěním byl ostrov v literatuře o drogových závislostech ignorován, snad proto, že není známo, že by byl přímým cílem mezokortikálního dopamin systém, který je ústředním bodem současných teorií závislosti na odměňování dopaminem. Výzkum publikovaný v roce 2007[76] to ukázal cigareta kuřáci trpící poškozením ostrovní kůry, od a mrtvice například nechte jejich závislost na cigaretách prakticky vyloučit. Bylo zjištěno, že u těchto jedinců je až 136krát vyšší pravděpodobnost, že dojde k narušení závislosti na kouření, než u kuřáků s poškozením v jiných oblastech. Narušení závislosti bylo prokázáno změnami chování hlášenými samy, jako je odvykání kouření méně než jeden den po poranění mozku, odvykání kouření s velkou lehkostí, nekouření po odvykání znovu a nutkání pokračovat v kouření od odvykání. Studie byla provedena v průměru osm let po mrtvici, což otevírá možnost, že připomenout zaujatost mohlo ovlivnit výsledky.[77] Novější prospektivní studie, které toto omezení překonaly, potvrdily tato zjištění[78][79] To naznačuje významnou roli ostrovní kůry v neurologických mechanismech, na nichž je závislost závislá nikotin a další léky, a učinil z této oblasti mozku možný cíl pro nové léky proti závislostem. Kromě toho toto zjištění naznačuje, že funkce zprostředkované ostrovem, zejména pocity vědomí, mohou být pro udržení drogové závislosti obzvláště důležité, i když tento pohled není zastoupen v žádném moderním výzkumu nebo přehledech subjektu.[80]
Nedávná studie na potkanech, kterou provedli Contreras et al.[81] potvrzuje tato zjištění tím, že ukazuje, že reverzibilní inaktivace izolátu narušuje amfetamin preferované podmíněné místo, zvířecí model toužení po drogách vyvolaný narážkou. V této studii inaktivace izolace také narušila reakce „malátnosti“ na chlorid lithný injekce, což naznačuje, že zastoupení negativních interoceptivních stavů ostrovem hraje roli v závislosti. V téže studii však došlo k preferenci podmíněného místa bezprostředně po injekci amfetaminu, což naznačuje, že na ostrově jsou zastoupeny okamžité, příjemné interoceptivní účinky podávání amfetaminu, spíše než opožděné averzní účinky stažení amfetaminu. .
Model navržený Naqvi et al. (viz výše) spočívá v tom, že na ostrově je uložena reprezentace příjemných interoceptivních účinků užívání drog (např. senzorické účinky nikotinu na dýchací cesty, kardiovaskulární účinky amfetaminu) a že toto vyjádření je aktivováno vystavením narážkám, které byly dříve spojené s užíváním drog. Řada funkčních zobrazovacích studií ukázala, že se izolace aktivuje během podávání návykových psychoaktivních drog. Několik funkčních zobrazovacích studií také ukázalo, že ostrov je aktivován, když jsou uživatelé drog vystaveni narážkám na drogy, a že tato aktivita koreluje se subjektivními nutkáními. Ve studiích cue-expozice je aktivita izolace vyvolána, pokud nedojde ke skutečné změně hladiny léku v těle. Proto může ostrov spíše než jen reprezentovat interoceptivní účinky užívání drog, jak k nim dochází, může hrát roli v paměti pro příjemné interoceptivní účinky užívání drog v minulosti, předvídání těchto účinků v budoucnosti nebo obojí. Taková reprezentace může vést k vědomým nutkáním, které mají pocit, jako by vycházely z těla. To může mít závislí pocit, jako by jejich těla potřebovala užívat drogu, a může to vést k tomu, že osoby s lézemi na ostrově hlásí, že jejich těla zapomněla na nutkání užívat, podle této studie.
Subjektivní jistota u extatických záchvatů
Běžnou vlastností mystických zážitků je silný pocit jistoty nelze vyjádřit slovy. Fabienne Picard navrhuje neurologické vysvětlení této subjektivní jistoty na základě klinického výzkumu epilepsie.[82][83] Podle Picarda může být tento pocit jistoty způsoben dysfunkcí přední insula, části mozku, která se podílí na interoception, sebereflexe a předcházení nejistotě ohledně vnitřních reprezentací světa „očekáváním řešení nejistoty nebo rizika“. Toto vyhýbání se nejistotě funguje porovnáním předpovězených a skutečných stavů, tj. „Signalizujících, že nerozumíme, tj. Že existuje nejednoznačnost“.[84] Picard poznamenává, že „koncept vhledu je velmi blízký konceptu jistoty“, a odkazuje na Archimédův „Heuréka!“[85][86] Picard předpokládá, že během extatických záchvatů již nefunguje srovnání mezi předpovězeným stavem a skutečným stavem a že již nebudou zpracovávány nesoulady mezi předpovězeným stavem a skutečným stavem, což blokuje „negativní emoce a negativní vzrušení vyplývající z prediktivní nejistoty“, které budou vnímány jako emoční sebevědomí.[87] Picard dochází k závěru, že „u některých jedinců by mohl vést k duchovní interpretaci“.[87]
Jiné klinické stavy
Bylo zjištěno, že ostrovní kůra má roli v úzkostných poruchách,[88] emoční dysregulace,[89] a mentální anorexie.[90]
Dějiny
Ostrov poprvé popsal Johann Christian Reil při popisu kraniálních a spinálních nervů a plexusů.[91] Henry Gray v Grayova anatomie je zodpovědný za to, že je známý jako Ostrov Reil.[91] John Allman a kolegové ukázali, že přední ostrovní kůra obsahuje vřetenové neurony.
Další obrázky
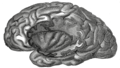
Izolace levé strany, odkrytá odstraněním opercula.
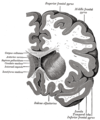
Koronální řez předními rohy laterálních komor.

Horizontální část levé mozkové hemisféry.



Viz také
Reference
- ^ MUFSON, E; MESULAM, M; PANDYA, D (1. července 1981). "Ostrovní propojení s amygdalou v opici rhesus". Neurovědy. 6 (7): 1231–1248. doi:10.1016/0306-4522(81)90184-6. PMID 6167896. S2CID 46366616.
- ^ Craig AD, Chen K, Bandy D, Reiman EM (2000). "Termosenzorická aktivace ostrovní kůry". Nat. Neurosci. 3 (2): 184–90. doi:10.1038/72131. PMID 10649575. S2CID 7077496.
- ^ JAKAB, A; MOLNAR, P; BOGNER, P; BERES, M; BERENYI, E (1. října 2011). „Připojení založené na propojení odhaluje interhemisférické rozdíly na ostrově“. Topografie mozku. 25 (3): 264–271. doi:10.1007 / s10548-011-0205-r. PMID 22002490. S2CID 12293575.
- ^ A b C Johannes Sobotta. „Sobottův Atlas a učebnice lidské anatomie 1909“. p. 145. Citováno 10. listopadu 2013.
- ^ „Definice:‚ Kruhový Sulcus z ostrova Insula'". MediLexicon. Citováno 2012-03-30.
- ^ Bauernfeind A; et al. (Duben 2013). „Objemové srovnání ostrovní kůry a jejích podoblastí u primátů“. Lidská evoluce. 64 (4): 263–279. doi:10.1016 / j.jhevol.2012.12.003. PMC 3756831. PMID 23466178.<
- ^ Mozek, MSN Encarta. Archivováno 2009-10-31.
- ^ Kolb, Bryan; Whishaw, Ian Q. (2003). Základy lidské neuropsychologie (5. vydání). [New York]: Stojí za to. ISBN 978-0-7167-5300-1.
- ^ Bushara, KO; Grafman, J; Hallett, M (1. ledna 2001). "Neurální koreláty detekce sluchově-vizuálního stimulu asynchronní detekce". The Journal of Neuroscience. 21 (1): 300–4. doi:10.1523 / JNEUROSCI.21-01-00300.2001. PMC 6762435. PMID 11150347.
- ^ Bushara, KO; Hanakawa, T; Immisch, I; Toma, K; Kansaku, K; Hallett, M (únor 2003). "Neurální koreláty křížově modální vazby". Přírodní neurovědy. 6 (2): 190–5. doi:10.1038 / nn993. PMID 12496761. S2CID 1098979.
- ^ Marieb, Elaine N .; Hoehn, Katja (2008). Anatomy & Physiology, Third Edition. Boston: Benjamin Cummings / Pearson. 391–395. ISBN 978-0-8053-0094-9.
- ^ Pritchard, TC; Macaluso, DA; Eslinger, PJ (srpen 1999). "Chuťové vnímání u pacientů s lézemi ostrovní kůry". Behaviorální neurovědy. 113 (4): 663–71. doi:10.1037/0735-7044.113.4.663. PMID 10495075.
- ^ Benedetto De Martino; Dharshan Kumaran; Ben Seymour; Raymond J. Dolan (srpen 2006). „Rámečky, předsudky a racionální rozhodování v lidském mozku“. Věda. 313 (6): 684–687. Bibcode:2006Sci ... 313..684D. doi:10.1126 / science.1128356. PMC 2631940. PMID 16888142.
- ^ Gui Xue; Zhonglin Lu; Irwin P. Levin d; Antoine Bechara (2010). „Dopad předchozích zkušeností s rizikem na následné riskantní rozhodování: Role ostrova“. NeuroImage. 50 (2): 709–716. doi:10.1016 / j.neuroimage.2009.12.097. PMC 2828040. PMID 20045470.
- ^ Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ (únor 2004). "Neuronové systémy podporující interoceptivní vědomí". Nat. Neurosci. 7 (2): 189–95. doi:10.1038 / nn1176. hdl:21.11116 / 0000-0001-A2FB-D. PMID 14730305. S2CID 13344271.
- ^ A b Lamb K, Gallagher K, McColl R, Mathews D, Querry R, Williamson JW (duben 2007). „Cvičením indukovaný pokles rCBF v ostrovní kůře během hypotenze po cvičení“. Med Sci Sports Exerc. 39 (4): 672–9. doi:10,1249 / mss.0b013e31802f04e0. PMID 17414805.
- ^ Williamson JW, McColl R, Mathews D, Mitchell JH, Raven PB, Morgan WP (duben 2001). „Hypnotická manipulace se smyslem pro úsilí během dynamického cvičení: kardiovaskulární reakce a aktivace mozku“. J. Appl. Physiol. 90 (4): 1392–9. doi:10.1152 / jappl.2001.90.4.1392. PMID 11247939. S2CID 8653997.
- ^ Williamson JW, McColl R, Mathews D, Ginsburg M, Mitchell JH (září 1999). „Aktivace ostrovní kůry je ovlivněna intenzitou cvičení“. J. Appl. Physiol. 87 (3): 1213–9. CiteSeerX 10.1.1.492.2730. doi:10.1152 / jappl.1999.87.3.1213. PMID 10484598.
- ^ Baliki MN, Geha PY, Apkarian AV (únor 2009). „Analýza vnímání bolesti mezi nociceptivní reprezentací a odhadem velikosti“. J. Neurophysiol. 101 (2): 875–87. doi:10.1152 / jn. 91100.2008. PMC 3815214. PMID 19073802.
- ^ Ogino Y, Nemoto H, Inui K, Saito S, Kakigi R, Goto F (květen 2007). „Vnitřní zkušenost s bolestí: představivost bolesti při sledování obrazů zobrazujících bolestivé události vytváří subjektivní představu bolesti v lidském mozku“. Cereb. Kůra. 17 (5): 1139–46. doi:10.1093 / cercor / bhl023. PMID 16855007.
- ^ Song GH, Venkatraman V, Ho KY, Chee MW, Yeoh KG, Wilder-Smith CH (prosinec 2006). „Kortikální účinky očekávání a endogenní modulace viscerální bolesti hodnocené funkční mozkovou MRI u pacientů se syndromem dráždivého tračníku a zdravých kontrol“. Bolest. 126 (1–3): 79–90. doi:10.1016 / j.pain.2006.06.017. PMID 16846694. S2CID 21437784.
- ^ Olausson H, Charron J, Marchand S, Villemure C, Strigo IA, Bushnell MC (listopad 2005). "Pocity tepla korelují s nervovou aktivitou v pravé přední ostrovní kůře". Neurosci. Lett. 389 (1): 1–5. doi:10.1016 / j.neulet.2005.06.065. PMID 16051437. S2CID 20068852.
- ^ Craig AD, Chen K, Bandy D, Reiman EM (únor 2000). "Termosenzorická aktivace ostrovní kůry". Nat. Neurosci. 3 (2): 184–90. doi:10.1038/72131. PMID 10649575. S2CID 7077496.
- ^ Ladabaum U, Minoshima S, Hasler WL, Cross D, Chey WD, Owyang C (únor 2001). "Distribuce žaludku koreluje s aktivací více kortikálních a subkortikálních oblastí". Gastroenterologie. 120 (2): 369–76. doi:10.1053 / gast.2001.21201. PMID 11159877.
- ^ Hamaguchi T, Kano M, Rikimaru H a kol. (Červen 2004). "Aktivita mozku během distenze sestupného tračníku u lidí". Neurogastroenterol. Motil. 16 (3): 299–309. doi:10.1111 / j.1365-2982.2004.00498.x. PMID 15198652.[mrtvý odkaz ]
- ^ Matsuura S, Kakizaki H, Mitsui T, Shiga T, Tamaki N, Koyanagi T (listopad 2002). „Reakce oblasti lidského mozku na distenzi nebo stimulaci chladu močového měchýře: studie pozitronové emisní tomografie“. J. Urol. 168 (5): 2035–9. doi:10.1016 / s0022-5347 (05) 64290-5. PMID 12394703.
- ^ von Leupoldt, A .; Sommer, T .; Kegat, S .; Baumann, H. J .; Klose, H .; Dahme, B .; Buchel, C. (24. ledna 2008). „Nepříjemnost vnímané dušnosti se zpracovává v přední části ostrova Insula a Amygdala“. American Journal of Respiratory and Critical Care Medicine. 177 (9): 1026–1032. doi:10.1164 / rccm.200712-1821OC. PMID 18263796.[trvalý mrtvý odkaz ]
- ^ Kikuchi M, Naito Y, Senda M a kol. (Duben 2009). "Kortikální aktivace během optokinetické stimulace - studie fMRI". Acta Otolaryngol. 129 (4): 440–3. doi:10.1080/00016480802610226. PMID 19116795. S2CID 42990194.
- ^ Papathanasiou ES, Papacostas SS, Charalambous M, Eracleous E, Thodi C, Pantzaris M (2006). "Závratě a nerovnováha způsobená malou lézí v přední izolaci". Electromyogr Clin Neurophysiol. 46 (3): 185–92. PMID 16918202.
- ^ Brown S, Martinez MJ, Parsons LM (září 2004). "Pasivní poslech hudby spontánně zapojuje limbické a paralimbické systémy". NeuroReport. 15 (13): 2033–7. doi:10.1097/00001756-200409150-00008. PMID 15486477. S2CID 12308683.
- ^ Sander K, Scheich H (říjen 2005). „Levá sluchová kůra a amygdala, ale pravá dominance ostrova pro lidský smích a pláč“. J. Cogn Neurosci. 17 (10): 1519–31. doi:10.1162/089892905774597227. PMID 16269094. S2CID 9509954.
- ^ „Archivovaná kopie“. Archivovány od originál dne 14. 7. 2010. Citováno 2010-07-04.CS1 maint: archivovaná kopie jako titul (odkaz)
- ^ Bamiou DE, Musiek FE, Luxon LM (květen 2003). "Ostrov (ostrov Reil) a jeho role ve sluchovém zpracování. Přehled literatury". Brain Res. Brain Res. Rev. 42 (2): 143–54. doi:10.1016 / S0165-0173 (03) 00172-3. PMID 12738055. S2CID 22339177.
- ^ Anderson TJ, Jenkins IH, Brooks DJ, Hawken MB, Frackowiak RS, Kennard C (říjen 1994). „Kortikální kontrola sakád a fixace u člověka. Studie PET“. Mozek. 117 (Pt 5): 1073–84. doi:10.1093 / mozek / 117.5.1073. PMID 7953589.
- ^ Fink GR, Frackowiak RS, Pietrzyk U, Passingham RE (duben 1997). „Několik neproměnných motorických oblastí v lidské kůře“. J. Neurophysiol. 77 (4): 2164–74. doi:10.1152 / jn.1997.77.4.2164. PMID 9114263. S2CID 15881491.
- ^ Sörös P, Inamoto Y, Martin RE (srpen 2009). „Funkční zobrazování mozku při polykání: metaanalýza odhadu pravděpodobnosti aktivace“. Hum Brain Mapp. 30 (8): 2426–39. doi:10,1002 / hbm.20680. PMC 6871071. PMID 19107749. S2CID 15438676.
- ^ Penfield W, Faulk ME (1955). „Ostrov; další pozorování jeho funkce“. Mozek. 78 (4): 445–70. doi:10.1093 / mozek / 78.4.445. PMID 13293263.
- ^ Dronkers NF (listopad 1996). "Nová oblast mozku pro koordinaci artikulace řeči". Příroda. 384 (6605): 159–61. Bibcode:1996 Natur.384..159D. doi:10.1038 / 384159a0. PMID 8906789. S2CID 4305696.
- ^ Ackermann H, Riecker A (květen 2004). „Příspěvek ostrova k motorickým aspektům produkce řeči: přehled a hypotéza“. Brain Lang. 89 (2): 320–8. doi:10.1016 / S0093-934X (03) 00347-X. PMID 15068914. S2CID 36867434.
- ^ Nowak M, Holm S, Biering-Sørensen F, Secher NH, Friberg L (červen 2005). ""Centrální povel „a ostrovní aktivace během pokusu o zvedání nohou u paraplegických lidí“. Hum Brain Mapp. 25 (2): 259–65. doi:10,1002 / hbm.20097. PMC 6871668. PMID 15849712.
- ^ Borovsky A, Saygin AP, Bates E, Dronkers N (červen 2007). „Léze koreluje s deficity produkce konverzační řeči“. Neuropsychologie. 45 (11): 2525–33. doi:10.1016 / j.neuropsychologia.2007.03.023. PMC 5610916. PMID 17499317.
- ^ Mutschler I, Schulze-Bonhage A, Glauche V, Demandt E, Speck O, Ball T (2007). Fitch T (ed.). „Rychlý zvukově-akční asociační efekt v lidské ostrovní kůře“. PLOS ONE. 2 (2): e259. Bibcode:2007PLoSO ... 2..259M. doi:10.1371 / journal.pone.0000259. PMC 1800344. PMID 17327919.
- ^ Weiller C, Ramsay SC, Wise RJ, Friston KJ, Frackowiak RS (únor 1993). "Jednotlivé vzorce funkční reorganizace v lidské mozkové kůře po kapsulárním infarktu". Annals of Neurology. 33 (2): 181–9. doi:10,1002 / ana 410330208. PMID 8434880.
- ^ Oppenheimer SM, Gelb A, Girvin JP, Hachinski VC (září 1992). "Kardiovaskulární účinky stimulace lidské ostrovní kůry". Neurologie. 42 (9): 1727–32. doi:10.1212 / mn.42.9.1727. PMID 1513461. S2CID 32371468.
- ^ Critchley HD (prosinec 2005). "Nervové mechanismy autonomní, afektivní a kognitivní integrace". J. Comp. Neurol. 493 (1): 154–66. doi:10.1002 / kně.20749. PMID 16254997.
- ^ Pacheco-López G, Niemi MB, Kou W, Härting M, Fandrey J, Schedlowski M (březen 2005). "Neurální substráty pro behaviorálně podmíněnou imunosupresi u potkanů". J. Neurosci. 25 (9): 2330–7. doi:10.1523 / JNEUROSCI.4230-04.2005. PMC 6726099. PMID 15745959.
- ^ Ramírez-Amaya V, Alvarez-Borda B, Ormsby CE, Martínez RD, Pérez-Montfort R, Bermúdez-Rattoni F (červen 1996). „Léze ostrovního kortexu zhoršují získávání podmíněné imunosuprese“. Brain Behav. Immun. 10 (2): 103–14. doi:10.1006 / brbi.1996.0011. PMID 8811934. S2CID 24813018.
- ^ Ramírez-Amaya V, Bermúdez-Rattoni F (březen 1999). „Podmíněné zvýšení produkce protilátek je narušeno ostrovní kůrou a amygdalou, ale ne hipokampálními lézemi“. Brain Behav. Immun. 13 (1): 46–60. doi:10.1006 / brbi.1998.0547. PMID 10371677. S2CID 20527835.
- ^ Karnath HO, Baier B, Nägele T (srpen 2005). „Povědomí o fungování vlastních končetin zprostředkované ostrovní kůrou?“. J. Neurosci. 25 (31): 7134–8. doi:10.1523 / JNEUROSCI.1590-05.2005. PMC 6725240. PMID 16079395.
- ^ Craig AD (leden 2009). „Jak se cítíš - teď? Přední ostrov a lidské vědomí“. Recenze přírody Neurovědy. 10 (1): 59–70. doi:10.1038 / nrn2555. PMID 19096369. S2CID 2340032.
- ^ Farrer C, Frith CD (březen 2002). „Prožívání sebe sama s jinou osobou jako příčiny činu: neurální koreláty zkušenosti agentury“. NeuroImage. 15 (3): 596–603. doi:10.1006 / nimg.2001.1009. PMID 11848702. S2CID 768408.
- ^ Tsakiris M, Hesse MD, chlapec C, Haggard P, Fink GR (říjen 2007). „Neurální podpisy vlastnictví těla: senzorická síť pro tělesné vědomí sebe sama“. Cereb. Kůra. 17 (10): 2235–44. doi:10.1093 / cercor / bhl131. PMID 17138596.
- ^ Wicker B, Keysers C, Plailly J, Royet JP, Gallese V, Rizzolatti G (říjen 2003). „Oba jsme znechuceni na Mé ostrově: společný nervový základ znechucení vidění a pocitu“. Neuron. 40 (3): 655–64. doi:10.1016 / S0896-6273 (03) 00679-2. PMID 14642287. S2CID 766157.
- ^ Wright P, He G, Shapira NA, Goodman WK, Liu Y (říjen 2004). "Znechucení a ostrov: reakce fMRI na obrázky zmrzačení a kontaminace". NeuroReport. 15 (15): 2347–51. doi:10.1097/00001756-200410250-00009. PMID 15640753. S2CID 6864309.
- ^ Jabbi M, Bastiaansen J, Keysers C (2008). Lauwereyns J (ed.). „Společná přední izolační reprezentace znechucení, zkušeností a představivosti ukazuje odlišné funkční cesty propojení“. PLOS ONE. 3 (8): e2939. Bibcode:2008PLoSO ... 3.2939J. doi:10,1371 / journal.pone 0002939. PMC 2491556. PMID 18698355.
- ^ Sanfey AG, Rilling JK, Aronson JA, Nystrom LE, Cohen JD (červen 2003). „Nervový základ ekonomického rozhodování ve hře Ultimatum“. Věda. 300 (5626): 1755–8. Bibcode:2003Sci ... 300.1755S. doi:10.1126 / science.1082976. PMID 12805551. S2CID 7111382.
- ^ Phan KL, Wager T, Taylor SF, Liberzon I (červen 2002). „Funkční neuroanatomie emocí: metaanalýza studií aktivace emocí u PET a fMRI“. NeuroImage. 16 (2): 331–48. doi:10.1006 / nimg.2002.1087. PMID 12030820. S2CID 7150871.
- ^ Singer T (2006). „Neuronální základ a ontogeneze empatie a čtení mysli: přehled literatury a důsledky pro budoucí výzkum“. Neurosci Biobehav Rev. 30 (6): 855–63. doi:10.1016 / j.neubiorev.2006.06.011. PMID 16904182. S2CID 15411628.
- ^ Ortigue S, Grafton ST, Bianchi-Demicheli F (srpen 2007). „Korelace mezi aktivací ostrova a kvalitou orgasmu u žen. NeuroImage. 37 (2): 551–60. doi:10.1016 / j.neuroimage.2007.05.026. PMID 17601749. S2CID 3377994.
- ^ Quarto, Tiziana; Blasi, Giuseppe; Maddalena, Chiara; Viscanti, Giovanna; Lanciano, Tiziana; Soleti, Emanuela; Mangiulli, Ivan; Taurisano, Paolo; Fazio, Leonardo (02.02.2016). „Sdružení mezi schopností emoční inteligence schopností a levou ostrovem během sociálního úsudku emocí obličeje“. PLOS ONE. 11 (2): e0148621. Bibcode:2016PLoSO..1148621Q. doi:10.1371 / journal.pone.0148621. ISSN 1932-6203. PMC 4747486. PMID 26859495.
- ^ Sázka, Tor (červen 2002). „Funkční neuroanatomie emocí: metaanalýza studií aktivace emocí u PET a fMRI“. NeuroImage. 16 (2): 331–48. doi:10.1006 / nimg.2002.1087. PMID 12030820. S2CID 7150871.
- ^ Vilares I, Howard JD, Fernandes HL, Gottfried JA, Kording KP (2012). „Diferenciální reprezentace předchozí a pravděpodobné nejistoty v lidském mozku“. Aktuální biologie. 22 (18): 1641–1648. doi:10.1016 / j.cub.2012.07.010. PMC 3461114. PMID 22840519.
- ^ Craig, A. D. (Bud) (2009). „Jak se cítíš - teď? Přední ostrov a lidské vědomí“ (PDF). Recenze přírody Neurovědy. 10 (1): 59–70. doi:10.1038 / nrn2555. PMID 19096369. S2CID 2340032. Archivovány od originál (PDF) dne 01.01.2013.
- ^ Craig, A. D. (Bud) (2002). „Nový pohled na bolest jako homeostatickou emoci“ (PDF). Trendy v neurovědách. 26 (6): 303–307. doi:10.1016 / s0166-2236 (03) 00123-1. PMID 12798599. S2CID 19794544. Archivovány od originál (PDF) dne 22.06.2010. Citováno 2009-09-03.
- ^ Lazar SW, Kerr CE, Wasserman RH, Gray JR, Greve DN, Treadway MT, McGarvey M, Quinn BT, Dusek JA, Benson H, Rauch SL, Moore CI, Fischl B (2005). „Meditace je spojena se zvýšenou tloušťkou kortikální oblasti“. NeuroReport. 16 (17): 1893–7. doi:10.1097 / 01.wnr.0000186598.66243.19. PMC 1361002. PMID 16272874.
- ^ Fox, Kieran C.R .; Nijeboer, Savannah; Dixon, Matthew L .; Floman, James L .; Ellamil, Melissa; Rumak, Samuel P .; Sedlmeier, Peter; Christoff, Kalina (červen 2014). „Je meditace spojena se změněnou strukturou mozku? Systematický přehled a metaanalýza morfometrického neuroobrazu u lékařů“. Neurovědy a biobehaviorální recenze. 43: 48–73. doi:10.1016 / j.neubiorev.2014.03.016. PMID 24705269. S2CID 207090878.
- ^ Hölzel, Britta K .; Ott, Ulrich; Gard, Tim; Hempel, Hannes; Weygandt, Martin; Morgen, Katrin; Vaitl, Dieter (2008). „Vyšetřování odborníků na meditaci všímavosti pomocí voxelové morfometrie“. Sociální kognitivní a afektivní neurovědy. 3 (1): 55–61. doi:10.1093 / sken / nsm038. PMC 2569815. PMID 19015095.
- ^ Damasio, A .; Damasio, H .; Tranel, D. (2013). „Přetrvávání pocitů a vnímání po oboustranném poškození ostrova“. Mozková kůra. 23 (4): 833–846. doi:10.1093 / cercor / bhs077. PMC 3657385. PMID 22473895.
- ^ Taylor KS, Seminowicz DA, Davis KD (září 2009). „Dva systémy spojení v klidovém stavu mezi kůrou insula a cingulate“. Hum Brain Mapp. 30 (9): 2731–45. doi:10,1002 / hbm.20705. PMC 6871122. PMID 19072897. S2CID 12917288.
- ^ A b Eckert MA, Menon V, Walczak A, Ahlstrom J, Denslow S, Horwitz A, Dubno JR (2009). „V srdci systému ventrální pozornosti: pravá přední izolace“. Hučení. Brain Mapp. 30 (8): 2530–41. doi:10,1002 / hbm.20688. PMC 2712290. PMID 19072895.
- ^ A b Zhang, Yang; Zhou, Wenjing; Wang, Siyu; Zhou, Qin; Wang, Haixiang; Zhang, Bingqing; Huang, Juan; Hong, Bo; Wang, Xiaoqin (01.02.2019). „Role členění lidské ostrovní oblasti na vnímání emocí a sluchové zpracování“. Mozková kůra. 29 (2): 517–528. doi:10.1093 / cercor / bhx334. ISSN 1047-3211. PMID 29342237. S2CID 36927038.
- ^ Blenkmann, Alejandro O .; Collavini, Santiago; Lubell, James; Llorens, Anaïs; Funderud, Ingrid; Ivanovic, Jugoslav; Larsson, Pål G .; Meling, Torstein R .; Bekinschtein, Tristan; Kochen, Silvia; Endestad, Tor (prosinec 2019). „Detekce sluchové odchylky na lidské ostrově: intrakraniální studie EEG“. Kůra. 121: 189–200. doi:10.1016 / j.cortex.2019.09.002. PMID 31629197. S2CID 202749677.
- ^ Afif, Afif; Minotti, Lorella; Kahane, Philippe; Hoffmann, Dominique (listopad 2010). „Anatomofunkční organizace ostrovní kůry: Studie využívající intracerebrální elektrickou stimulaci u epileptických pacientů: Funkční organizace ostrova Insula“. Epilepsie. 51 (11): 2305–2315. doi:10.1111 / j.1528-1167.2010.02755.x. PMID 20946128.
- ^ Nestor PJ, Graham NL, Fryer TD, Williams GB, Patterson K, Hodges JR (listopad 2003). „Progresivní neplynulá afázie je spojena s hypometabolismem soustředěným na levé přední insula“. Mozek. 126 (Pt 11): 2406–18. doi:10.1093 / mozek / awg240. PMID 12902311.
- ^ Gorno-Tempini ML, Dronkers NF, Rankin KP a kol. (Březen 2004). „Poznání a anatomie ve třech variantách primární progresivní afázie“. Annals of Neurology. 55 (3): 335–46. doi:10.1002 / ana.10825. PMC 2362399. PMID 14991811.
- ^ Nasir H. Naqvi; David Rudrauf; Hanna Damasio; Antoine Bechara. (Leden 2007). „Poškození ostrova narušuje závislost na kouření cigaret“. Věda. 315 (5811): 531–4. Bibcode:2007Sci ... 315..531N. doi:10.1126 / science.1135926. PMC 3698854. PMID 17255515.
- ^ Vorel SR, Bisaga A, McKhann G, Kleber HD (červenec 2007). "Poškození izolace a ukončení kouření". Věda. 317 (5836): 318–9, odpověď autora 318–9. doi:10.1126 / science.317.5836.318c. PMID 17641181. S2CID 8917168.
- ^ Suner-Soler, R. (2011). „Odvykání kouření 1 rok po mrtvici a poškození ostrovní kůry“. Mrtvice. 43 (1): 131–136. doi:10.1161 / STROKEAHA.111.630004. PMID 22052507.
- ^ Gaznick, N. (2013). „Poškození na bazálech Ganglia Plus způsobí silnější narušení závislosti na kouření, než samotné poškození bazální ganglií“. Nikotin. 16 (4): 445–453. doi:10.1093 / ntr / ntt172. PMC 3954424. PMID 24169814.
- ^ Hyman, Steven E. (01.08.2005). „Závislost: Nemoc z učení a paměti“. Jsem J. Psychiatrie. 162 (8): 1414–22. doi:10.1176 / appi.ajp.162.8.1414. PMID 16055762.
- ^ Marco Contreras; Francisco Ceric; Fernando Torrealba (leden 2007). „Inaktivace interoceptivní látky Insula narušuje touhu po drogách a malátnost vyvolanou lithiem“. Věda. 318 (5850): 655–8. Bibcode:2007Sci ... 318..655C. doi:10.1126 / science.1145590. PMID 17962567. S2CID 23499558.
- ^ Picard, Fabienne (2013), „Stav víry, subjektivní jistoty a blaženosti jako produkt kortikální dysfunkce“, Kůra, 49 (9): 2494–2500, doi:10.1016 / j.cortex.2013.01.006, PMID 23415878, S2CID 206984751
- ^ Gschwind, Markus; Picard, Fabienne (2016), „Extatické epileptické záchvaty: letmý pohled na rozmanité role ostrova“, Hranice v behaviorální neurovědě, 10: 21, doi:10.3389 / fnbeh.2016.00021, PMC 4756129, PMID 26924970
- ^ Picard 2013, s. 2496-2498
- ^ Picard 2013, s. 2497-2498
- ^ Viz také satori v japonském zenu
- ^ A b Picard 2013, s. 2498
- ^ Paulus MP, Stein MB (August 2006). "Ostrovní pohled na úzkost". Biol. Psychiatrie. 60 (4): 383–7. doi:10.1016 / j.biopsych.2006.03.042. PMID 16780813. S2CID 17889111.
- ^ Thayer JF, Lane RD (December 2000). "A model of neurovisceral integration in emotion regulation and dysregulation". J Affect Disord. 61 (3): 201–16. doi:10.1016/S0165-0327(00)00338-4. PMID 11163422.
- ^ Gaudio S, Wiemerslage L, Brooks SJ, Schiöth HB (2016). „Systematický přehled studií funkční-MRI v klidovém stavu u mentální anorexie: Důkazy o zhoršení funkční konektivity v kognitivní kontrole a integraci visuospatial a body-signal“ (PDF). Neurosci Biobehav Rev. 71: 578–589. doi:10.1016 / j.neubiorev.2016.09.032. PMID 27725172. S2CID 16526824.
- ^ A b Binder DK, Schaller K, Clusmann H (November 2007). "The seminal contributions of Johann-Christian Reil to anatomy, physiology, and psychiatry". Neurochirurgie. 61 (5): 1091–6, discussion 1096. doi:10.1227/01.neu.0000303205.15489.23. PMID 18091285. S2CID 8152708.
externí odkazy
- Ostrovní kůra v Brede databáze na Technická univerzita v Dánsku. Location and literature citations for the insula
- synd/1212 na Kdo to pojmenoval?
- "Anatomický diagram: 13048 000-1". Roche Lexicon - ilustrovaný navigátor. Elsevier. Archivovány od originál dne 7. 11. 2014.
- Stained brain slice images which include the "insular cortex" na Projekt BrainMaps
- Thomas P. Naidicha; et al. (1. února 2004). "The Insula: Anatomic Study and MR Imaging Display at 1.5 T". American Journal of Neuroradiology. 25 (2): 222–32. PMID 14970021.
- Kakigia R, Nakataa H, Inuia K, Hiroea N, et al. (Říjen 2005). "Intracerebral pain processing in a Yoga Master who claims not to feel pain during meditation". Eur J Pain. 9 (5): 581–9. doi:10.1016/j.ejpain.2004.12.006. PMID 16139187.
As for fMRI recording, there were remarkable changes in levels of activity in the...SII-insula (mainly the insula)
| Kontrolní úřad |
|---|