Čichový neuron receptoru - Olfactory receptor neuron
| Čichový neuron receptoru | |
|---|---|
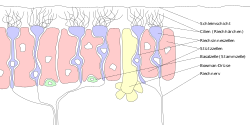 Štítky v němčině. "Zellen" = "buňka "," riech "=" vůně "," Riechnerv "= čichový nerv, "cillien" = řasy. | |
| Detaily | |
| Systém | Čich |
| Umístění | čichový epitel v nos |
| Tvar | Bipolární senzorický receptor |
| Funkce | Detekujte stopy chemikálií ve vdechovaném vzduchu (čich ) |
| Neurotransmiter | Glutamát[1] |
| Presynaptické spojení | Žádný |
| Postsynaptická spojení | Čichová žárovka |
| Identifikátory | |
| Pletivo | D018034 |
| NeuroLex ID | nifext_116 |
| TH | H3.11.07.0.01003 |
| Anatomické pojmy neuroanatomie | |
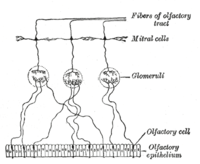

An čichový receptorový neuron (ORN), také nazývaný čichový senzorický neuron (OSN), je senzorický neuron v rámci čichový systém.[2]
Struktura
Lidé mají mezi 10 a 20 miliony neuronů čichových receptorů.[3] v obratlovců, ORN jsou bipolární neurony s dendrity čelem k vnějšímu povrchu cribriformní desky s axony, které procházejí foriem cribriform s koncovým koncem u čichových cibulí. ORN jsou umístěny v čichový epitel v nosní dutině. Buněčná těla ORN jsou rozdělena mezi všechny tři rozvrstvené vrstvy čichového epitelu.[4]
Mnoho drobných vlasů řasy vyčnívají z čichové receptorové buňky dendrit do sliz pokrývající povrch čichového epitelu. Povrch těchto řasinek je pokryt čichové receptory, typ Receptor spojený s G proteinem. Každá čichová receptorová buňka vyjadřuje pouze jeden typ čichového receptoru (OR), ale mnoho samostatných buněk čichového receptoru exprimuje OR, které váží stejnou sadu pachů. Axony čichových receptorových buněk, které exprimují stejný OR, se sbíhají do formy glomeruli v čichová žárovka.[5]
Funkce
OR, které se nacházejí na membránách řasinek, byly klasifikovány jako komplexní typ s ligandem metabotropní kanály.[6] Existuje přibližně 1000 různých genů, které kódují OR, což z nich dělá největší genovou rodinu. Odorant se rozpustí v hlenu čichového epitelu a poté se váže na OR. OR se mohou vázat na různé molekuly pachu s různou afinitou. Rozdíl v afinitách způsobuje rozdíly v aktivačních vzorcích, což vede k jedinečným profilům odorantů.[7][8] Aktivovaný OR zase aktivuje intracelulární G-protein, GOLF (GNAL ), adenylátcykláza a výroba cyklický AMP (cAMP) se otevře iontové kanály v buněčná membrána, což vedlo k přílivu sodík a vápník ionty do buňky a výtok chlorid ionty. Tento příliv kladných iontů a odtok záporných iontů způsobí depolarizaci neuronu a generování akční potenciál.
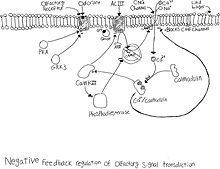
Desenzibilizace
Čichový receptorový neuron má rychle fungující negativní zpětnou vazbu depolarizace. Když je neuron depolarizující, Iontový kanál CNG je otevřený umožňující sodík a vápník spěchat do cely. Příliv vápníku začíná v buňce kaskádou událostí. Vápník se nejprve váže na kalmodulin a tvoří se Vačka. CaM se poté naváže na kanál CNG a uzavře jej, čímž zastaví přívod sodíku a vápníku.[9] CaMKII se aktivuje přítomností CaM, který fosforyluje ACIII a sníží produkci cAMP.[10] CaMKII se také aktivuje fosfodiesteráza, který poté hydrolyzuje cAMP.[11] Účinek této negativní zpětné vazby inhibuje neuron z další aktivace, když je zavedena další molekula zápachu.
Počet rozlišitelných pachů
Široce uveřejněná studie naznačuje, že lidé mohou detekovat více než jeden bilion různých pachů.[12] Toto zjištění však bylo zpochybněno. Kritici tvrdili, že metodika použitá pro odhad byla zásadně chybná, což ukazuje, že použití stejného argumentu pro lépe pochopené senzorické modality, jako je vize nebo konkurz, vede k nesprávným závěrům.[13] Jiní vědci také prokázali, že výsledek je extrémně citlivý na přesné detaily výpočtu, přičemž malé odchylky mění výsledek v řádu desítek řádů, případně až na několik tisíc.[14] Autoři původní studie tvrdili, že jejich odhad platí, pokud se předpokládá, že pachový prostor je dostatečně vysoce dimenzionální.[15]
Ostatní zvířata
Tato sekce potřebuje expanzi. Můžete pomoci přidávat k tomu. (Února 2018) |
Viz také
Reference
- ^ Berkowicz, D. A .; Trombley, P. Q .; Shepherd, G. M. (1994). "Důkazy pro glutamát jako neurotransmiter čichových receptorových buněk". Journal of Neurophysiology. 71 (6): 2557–61. doi:10.1152 / jn.1994.71.6.2557. PMID 7931535.
- ^ Vermeulen, A; Rospars, J. P. (1998). „Dendritická integrace v čichových senzorických neuronech: Analýza ustáleného stavu, jak struktura neuronů a prostředí neuronů ovlivňují kódování intenzity zápachu“. Journal of Computational Neuroscience. 5 (3): 243–66. doi:10.1023 / A: 1008826827728. PMID 9663551.
- ^ Saladin, Kenneth (2012). Anatomie a fyziologie: jednota formy a funkce (6. vydání). McGraw-Hill. str. 593. ISBN 978-0073378251.
- ^ Cunningham, A.M .; Manis, P.B .; Reed, R.R .; Ronnett, G.V. (1999). „Čichové receptorové neurony existují jako odlišné podtřídy nezralých a zralých buněk v primární kultuře“. Neurovědy. 93 (4): 1301–12. doi:10.1016 / s0306-4522 (99) 00193-1. PMID 10501454.
- ^ McEwen, D. P (2008). „Čichová řasinka: naše přímé neuronální spojení s vnějším světem“. Curr. Horní. Dev. Biol. Aktuální témata vývojové biologie. 85: 333–370. doi:10.1016 / S0070-2153 (08) 00812-0. ISBN 9780123744531. PMID 19147011.
- ^ Touhara, Kazushige (2009). „Složité funkce čichového receptoru hmyzu jako ionotropní kanál s ligandovou bránou“. Annals of the New York Academy of Sciences. 1170 (1): 177–80. Bibcode:2009NYASA1170..177T. doi:10.1111 / j.1749-6632.2009.03935.x. PMID 19686133.
- ^ Bieri, S .; Monastyrskaia, K; Schilling, B (2004). „Čichový receptor neuronového profilování s použitím odorantů santalového dřeva“. Chemické smysly. 29 (6): 483–7. doi:10.1093 / chemse / bjh050. PMID 15269120.
- ^ Fan, Jinhong; Ngai, John (2001). "Nástup genové exprese odorantového receptoru během regenerace čichového senzorického neuronu". Vývojová biologie. 229 (1): 119–27. doi:10,1006 / dbio.2000.9972. PMID 11133158.
- ^ Bradley, J; Reuter, D; Frings, S (2001). "Usnadnění adaptace pachu zprostředkovaného kalmodulinem pomocí kanálových podjednotek s bránou cAMP". Věda. 294 (5549): 2176–2178. Bibcode:2001Sci ... 294.2176B. doi:10.1126 / science.1063415. PMID 11739960.
- ^ Wei, J; Zhao, AZ; Chan, GC; Baker, LP; Impey, S; Beavo, JA; Storm, DR (1998). „Fosforylace a inhibice čichové adenylylcyklázy CaM kinázou II v neuronech: mechanismus pro útlum čichových signálů“. Neuron. 21 (3): 495–504. doi:10.1016 / s0896-6273 (00) 80561-9. PMID 9768837.
- ^ Yan, C; Zhao, AZ; Bentley, JK; Loughney, K; Ferguson, K; Beavo, JA (1995). „Molekulární klonování a charakterizace kalmodulin-dependentní fosfodiesterázy obohacené o čichové senzorické neurony“. Proc Natl Acad Sci USA. 92 (21): 9677–9681. Bibcode:1995PNAS ... 92,9677Y. doi:10.1073 / pnas.92.21.9677. PMC 40865. PMID 7568196.
- ^ Bushdid, C .; Magnasco, M. O .; Vosshall, L. B .; Keller, A. (2014). „Lidé mohou rozlišovat více než 1 bilion čichových stimulů“. Věda. 343 (6177): 1370–2. Bibcode:2014Sci ... 343.1370B. doi:10.1126 / science.1249168. PMC 4483192. PMID 24653035.
- ^ Meister, Markus (2015). „O rozměrnosti pachového prostoru“. eLife. 4: e07865. doi:10,7554 / eLife.07865. PMC 4491593. PMID 26151672.
- ^ Gerkin, Richard C .; Castro, Jason B. (2015). „Počet čichových podnětů, které mohou lidé rozlišovat, je stále neznámý“. eLife. 4: e08127. doi:10,7554 / eLife.08127. PMC 4491703. PMID 26151673.
- ^ Magnasco, Marcelo O .; Keller, Andreas; Vosshall, Leslie B. (2015). „O rozměrnosti čichového prostoru“. doi:10.1101/022103. Citovat deník vyžaduje
| deník =(Pomoc)
externí odkazy
| Kontrolní úřad |
|---|