Lidská taxonomie - Human taxonomy
| Homo („lidé“) | |
|---|---|
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Třída: | Mammalia |
| Objednat: | Primáti |
| Podřád: | Haplorhini |
| Infraorder: | Simiiformes |
| Rodina: | Hominidae |
| Podčeleď: | Homininae |
| Kmen: | Hominini |
| Rod: | Homo Linné, 1758 |
| Zadejte druh | |
| Homo sapiens Linné, 1758 | |
| Druh | |
navrhované jiné druhy nebo poddruhy | |
| Synonyma | |
Synonyma
| |
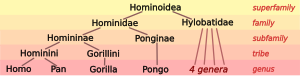
Lidská taxonomie je klasifikace lidský druh (systematické jméno Homo sapiens, latinský: "moudrý muž") v rámci zoologické taxonomie. Systematické rod, Homo, je navržen tak, aby zahrnoval obojí anatomicky moderní lidé a vyhynulé odrůdy archaičtí lidé. Současní lidé byli označeni jako poddruhy Homo sapiens sapiens, odlišené podle některých od přímého předka, Homo sapiens idaltu (s nějakým jiným výzkumem místo klasifikace idaltu a současní lidé patřící do stejného poddruhu[1][2][3]).
Od zavedení systematických názvů v 18. století, znalost lidská evoluce drasticky vzrostla a ve 20. až na počátku 21. století byla navržena řada přechodných taxonů. Nejuznávanější taxonomické skupiny tento rod přebírají Homo jako pocházející před dvěma až třemi miliony let, rozdělené na nejméně dva druhy, archaické Homo erectus a moderní Homo sapiens, s asi tuctem dalších návrhů na druhy bez univerzálního uznání.
Rod Homo je umístěn v kmen Hominini vedle Pánev (šimpanzi). Odhaduje se, že tyto dva rody mají rozcházely se po delší dobu hybridizace, která trvala zhruba před 10 až 6 miliony let, s možnou příměsí až před 4 miliony let. Pod kmen nejisté platnosti, seskupující archaické „předlidské“ nebo „parahumánní“ druhy mladší než Homo-Pánev split je Australopithecina (navrženo v roce 1939).
Návrh Wooda a Richmonda (2000) by představil Homininu jako pod kmen vedle Australopitheciny, s Homo jediný známý rod v rámci Homininy. Alternativně, po Cela-Conde a Ayale (2003), rody „předlidské“ nebo „proto-lidské“ Australopithecus, Ardipithecus, Praeanthropus, a možná Sahelanthropus mohou být postaveny na stejné úrovni vedle rodu Homo. Ještě radikálnější pohled odmítá rozdělení Pánev a Homo jako samostatné rody, které na základě Zásada priority by znamenalo přeřazení šimpanzů na Homo paniscus (nebo podobné).[4]
Před současnou vědeckou klasifikací lidí provedli filozofové a vědci různé pokusy o klasifikaci lidí. Nabízeli definice lidské bytosti a schémata pro klasifikaci typů lidí. Biologové kdysi klasifikovali rasy jako poddruhy, ale dnes antropologové odmítají koncept rasy a považují lidstvo za vzájemně související genetické kontinuum. Taxonomie homininů se nadále vyvíjí.[5][6]
Dějiny
Lidská taxonomie na jedné straně zahrnuje umístění lidí do Taxonomie USA hominidy (lidoopi), a na druhé straně rozdělení archaický a moderní člověk do druh a případně poddruh. Moderní zoologická taxonomie byla vyvinuta Carl Linné během třicátých a padesátých let. Pojmenoval lidský druh jako Homo sapiens v roce 1758 jako jediný člen rodu Homo, rozdělen na několik poddruhů odpovídající skvělé závody. The latinský podstatné jméno homo (genitiv hominis) znamená „člověk“. Systematický název Hominidae pro rodina velkých lidoopů představil John Edward Gray (1825).[7] Šedá také dodávána Hominini jako název kmen včetně obou šimpanzů (rod Pánev ) a lidé (rod Homo).
Objev prvního vyhynulého archaického lidského druhu z fosilních záznamů se datuje do poloviny 19. století, Homo neanderthalensis, klasifikovaný v roce 1864. Od té doby byla pojmenována řada dalších archaických druhů, ale neexistuje přesná shoda ohledně jejich přesného počtu. Po objevu H. neanderthalensis, který, i když je „archaický“ rozpoznatelný jako zjevně lidský, byla antropologie koncem 19. až začátkem 20. století po určitou dobu zaměstnána hledáním údajně “Chybějící odkaz „mezi Homo a Pánev. „Piltdown Man „Falešná zpráva z roku 1912 byla podvodná prezentace takového přechodného druhu. Od poloviny 20. století se znalosti vývoje Hominini staly mnohem podrobnějšími a taxonomická terminologie byla několikrát změněna, aby to odrážela.
Zavedení Australopithecus jako třetí rod, vedle Homo a Pánev, v kmeni Hominini je kvůli Raymond Dart (1925). Australopithecina jako pod kmen obsahující Australopithecus stejně jako Paranthropus (Koště 1938) je návrhem Gregoryho a Hellmana (1939). Nověji navržené dodatky k podkmenu Australopithecina zahrnují Ardipithecus (1995) a Kenyanthropus (2001). Pozice Sahelanthropus (2002) ve vztahu k Australopithecina v rámci Hominini je nejasný. Cela-Conde a Ayala (2003) navrhují uznání Australopithecus, Ardipithecus, Praeanthropus, a Sahelanthropus (dopis incertae sedis) jako samostatné rody.[8]
Další navrhované rody, nyní většinou považované za součást Homo, zahrnout:Pithecanthropus (Dubois, 1894),Protanthropus (Haeckel, 1895),Sinanthropus (Black, 1927),Cyphanthropus (Pycraft, 1928)Africanthropus (Dreyer, 1935),[9]Telanthropus (Broom & Anderson 1949),Atlanthropus (Arambourg, 1954),Tchadanthropus (Coppens, 1965).
Rod Homo byl vzat vzniknout asi před dvěma miliony let, od objevu kamenné nástroje v Olduvai Gorge, Tanzanie v 60. letech. Homo habilis (Leakey et al., 1964) by byl prvním „lidským“ druhem (člen rodu Homo) podle definice, přičemž jeho typovým exemplářem je OH 7 fosilie. Objev dalších fosilií tohoto typu však otevřel debatu o vymezení H. habilis z Australopithecus. Zvláště LD 350-1 fosilie čelistí objevená v roce 2013, datovaná na 2,8 Mya, byla považována za přechodnou mezi těmito dvěma.[10] Je rovněž sporné, zda H. habilis byl prvním homininem, který použil kamenné nástroje, jako Australopithecus garhi, datováno k c. 2.5 Mya, bylo nalezeno spolu s kamennými nástroji.[11] Fosilní KNM-ER 1470 (objeveno v roce 1972, označeno Pithecanthropus rudolfensis podle Alekseyev 1978) je nyní považován za třetí raný druh rodu Homo (vedle H. habilis a H. erectus) přibližně před 2 miliony let, nebo alternativně jako přechodné období mezi Australopithecus a Homo.[12]
Wood a Richmond (2000) navrhli Grayův kmen Hominini ("homininy") se označují jako zahrnující všechny druhy po šimpanz-člověk poslední společný předek podle definice k zařazení australopiteků a dalších možných předlidských nebo parahumánních druhů (jako např Ardipithecus a Sahelanthropus ) není známo v době Grayova.[13] V tomto návrhu měl být označen nový kmen kmene Hominina, který zahrnuje rod Homo výlučně proto, aby Hominini měl dvě podskupiny, Australopithecina a Hominina, přičemž jediný známý rod v Hominině byl Homo. Orrorin (2001) byl navržen jako možný předchůdce Homininy, nikoli však Australopitheciny.[14]
Byly navrženy alternativy k Hominině: Australopithecinae (Gregory & Hellman 1939) a Preanthropinae (Cela-Conde & Altaba 2002);[15]
Druh
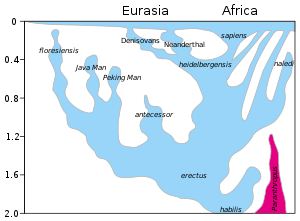
Nejméně tucet druhů Homo jiný než Homo sapiens byly navrženy s různou mírou shody. Homo erectus je široce uznáván jako druh přímo předků Homo sapiens.[Citace je zapotřebí ] Většina ostatních navrhovaných druhů se navrhuje jako alternativa k některému z nich Homo erectus nebo Homo sapiens jako poddruh. To se týká Homo ergaster zejména.[16][17] Jeden návrh rozděluje Homo erectus do africké a asijské odrůdy; Afričan je Homo ergaster, a ta asijská je Homo erectus sensu stricto. (Zahrnutí Homo ergaster s asijskými Homo erectus je Homo erectus sensu lato.)[18] Zdá se, že existuje nedávný trend s dostupností stále obtížněji klasifikovatelných fosilií, jako je Dmanisi lebky (2013) nebo Homo naledi fosílie (2015) k zařazení všech archaických odrůd pod Homo erectus.[19][20][21]
| Počty řádků | Časový rozsah kya | Místo výskytu | Výška dospělých | Hmotnost dospělých | Lebeční kapacita (cm³) | Fosilní záznam | Objevení / zveřejnění jména |
|---|---|---|---|---|---|---|---|
| H. habilis členství v Homo nejistý | 2,100–1,500[A][b] | Tanzanie | 110–140 cm (3 ft 7 in - 4 ft 7 in) | 33–55 kg (73–121 lb) | 510–660 | Mnoho | 1960 1964 |
| H. rudolfensis členství v Homo nejistý | 1,900 | Keňa | 700 | 2 stránky | 1972 1986 | ||
| H. gautengensis také klasifikován jako H. habilis | 1,900–600 | Jižní Afrika | 100 cm (3 ft 3 v) | 3 jednotlivci[24][C] | 2010 2010 | ||
| H. erectus | 1,900–140[25][d][26] Homo erectus soloensis, nalezen v Jáva, je považován za poslední známé přežití H. erectus. Studie z roku 2011, která byla dříve datována až před 50 000 až 40 000 lety, posunula datum jejího zániku H. e. soloensis nejpozději před 143 000 lety, pravděpodobněji před 550 000 lety. [27]}} | Afrika, Eurasie | 180 cm (5 ft 11 v) | 60 kg (130 lb) | 850 (brzy) - 1100 (pozdě) | Mnoho[E][F] | 1891 1892 |
| H. ergaster Afričan H. erectus | 1,800–1,300[28] | Východní a jižní Afrika | 700–850 | Mnoho | 1949 1975 | ||
| H. předchůdce | 1,200–800 | západní Evropa | 175 cm (5 ft 9 v) | 90 kg (200 lb) | 1,000 | 2 stránky | 1994 1997 |
| H. heidelbergensis brzy H. neanderthalensis | 600–300[G] | Evropa, Afrika | 180 cm (5 ft 11 v) | 90 kg (200 lb) | 1,100–1,400 | Mnoho | 1907 1908 |
| H. cepranensis možná jediná fosilie H. erectus | C. 450[29] | Itálie | 1,000 | 1 čepice lebky | 1994 2003 | ||
| H. rhodesiensis brzy H. sapiens | C. 300 | Zambie | 1,300 | svobodný nebo velmi málo | 1921 1921 | ||
| H. naledi | C. 300[30] | Jižní Afrika | 150 cm (4 ft 11 v) | 45 kg (99 lb) | 450 | 15 osob | 2013 2015 |
| H. sapiens (anatomicky moderní lidé) | C. 300 - přítomen[h] | Celosvětově | 150–190 cm (4 ft 11 in - 6 ft 3 in) | 50–100 kg (110–220 lb) | 950–1,800 | (existující) | —— 1758 |
| H. neanderthalensis | 240–40[33][i] | Evropa, západní Asie | 170 cm (5 ft 7 v) | 55–70 kg (121–154 lb) (těžce stavěné) | 1,200–1,900 | Mnoho | 1829 1864 |
| H. floresiensis klasifikace nejistá | 190–50 | Indonésie | 100 cm (3 ft 3 v) | 25 kg (55 lb) | 400 | 7 osob | 2003 2004 |
| H. tsaichangensis možná H. erectus | C. 100[j] | Tchaj-wan | 1 osoba | 2008(?) 2015 | |||
| H. luzonensis | C. 67[36][37] | Filipíny | 3 jednotlivci | 2007 2019 | |||
| Denisova hominin | 40 | Sibiř | 2 stránky | 2000 2010[k] | |||
| Jeskynní lidé možný H. sapiens poddruh nebo hybridní | 15–12[l][38] | Jihozápadní Čína | Velmi málo |
Poddruh
Homo sapiens poddruh

Uznání nebo neuznání poddruh z Homo sapiens má komplikovanou historii. Hodnost poddruhu v zoologii se zavádí pro pohodlí, a nikoli na základě objektivních kritérií založených na pragmatickém zvážení faktorů, jako jsou geografická izolace a sexuální výběr. Neformální taxonomická hodnost závod je různě považován za ekvivalent nebo podřízený hodnosti poddruhu a rozdělení anatomicky moderní lidé (H. sapiens) do poddruhu úzce souvisí s uznáním hlavní rasová uskupení na základě lidská genetická variace.
Poddruh nelze rozpoznat samostatně: druh bude uznán jako druh, který nemá vůbec žádný poddruh nebo alespoň dva (včetně těch, které již zanikly). Proto označení existujícího poddruhu Homo sapiens sapiens má smysl, pouze pokud je rozpoznán alespoň jeden další poddruh. H. S. sapiens je taxonomicky přičítán „Linné (1758)“ Zásada koordinace.[40] William Stearn (1959) v „absolvující poznámce“[41] tvrdil, že Linné „musí být typem jeho Homo sapiens“. Protože Linné popisuje H. s. europaeus jako modrá / zelená (caerulus) oči, ale sám měl hnědé oči, nemohl se do něj zahrnout H. s. europaeus, Linné by proto muselo být klasifikováno jako H. sapiens sapiens, protože neodpovídá žádnému z popisů jeho pěti poddruhů, a tak by stál jako lektotyp oba pro H. sapiens, a pro H. s. sapiens v rámci jeho vlastní poddruhové nomenklatury.[42]
Během 19. až poloviny 20. století bylo běžnou praxí klasifikovat hlavní divize existujících H. sapiens jako poddruh, po Linné (1758), který poznal H. s. americanus, H. s. europaeus, H. s. asiaticus a H. s. afer jako seskupení původních populací Amerika, Západní Eurasie, východní Asie a Subsaharská Afrika kromě toho H. s. ferus (pro „divokou“ formu, se kterou se ztotožnil divoké děti ) a dvě další „divoké“ formy pro nahlášené vzorky, které jsou nyní považovány za součást kryptozoologie, H. s. monstrosus a H. s. troglodyty.[43]
Byly provedeny variace a doplňky kategorií Linnaeus, jako např H. s. tasmanianus pro původní obyvatelstvo Austrálie.[44] Bory de St. Vincent v jeho Essai sur l'Homme (1825) rozšířil Linnérasový "kategorie až na patnáct: Leiotrichi ("hladké vlasy "): japeticus (s podchody), arabicus, indicus, scythicus, sinicus, hyperboreus, neptunianus, australasicus, columbicus, americanus, patagonicus; Oulotrichi ("křehké vlasy "): aethiopicus, cafer, hotentotus, melaninus.[45] Podobně, Georges Vacher de Lapouge (1899) měl také kategorie založené na rase, jako např priscus, spelaeus (atd.).
Homo sapiens neanderthalensis byl navržen Kingem (1864) jako alternativa k Homo neanderthalensis.[46] Od jejich objevu v šedesátých letech minulého století probíhaly „taxonomické války“ o to, zda byli neandertálci samostatným druhem. Pääbo (2014) to rámcuje jako debatu, která je v zásadě neřešitelná, „protože neexistuje žádná definice druhů, která by dokonale popisovala případ“.[47] Louis Lartet (1869) Homo sapiens fossilis založeno na Kromagnonské fosílie.
Existuje řada návrhů na vyhynulé odrůdy Homo sapiens vyrobené ve 20. století. Mnoho z původních návrhů nepoužívalo explicitní trinomiální nomenklatura, přestože jsou stále uváděny jako platné synonyma z H. sapiens Wilson & Reeder (2005).[48] Tyto zahrnují: Homo grimaldii (Lapouge, 1906),Homo aurignacensis hauseri (Klaatsch & Hauser, 1910),Notanthropus eurafricanus (Sergi, 1911), Homo fossilis infrasp. proto-aethiopicus (Giuffrida-Ruggeri, 1915),Telanthropus capensis (Broom, 1917),[49]Homo wadjakensis (Dubois, 1921), Homo sapiens cro-magnonensis, Homo sapiens grimaldiensis (Gregory, 1921),Homo drennani (Kleinschmidt, 1931),[50]Homo galilensis (Joleaud, 1931) = Paleanthropus palestinus (McCown a Keith, 1932).[51]Rightmire (1983) navrhl Homo sapiens rhodesiensis.[52]
Do 80. let 20. století byla praxe rozdělení existujících populací Homo sapiens do poddruhu klesal. Raná autorita, která se výslovně vyhýbá rozdělení H. sapiens do poddruhu byl Grzimeks Tierleben, publikováno 1967–1972.[53]Pozdní příklad akademické autority, která navrhuje, aby byly lidské rasové skupiny považovány za taxonomický poddruh, je John Baker (1974).[54] Trojčlenná nomenklatura Homo sapiens sapiens se stal populárním pro „moderní lidi“ v kontextu neandertálců, kteří byli považováni za poddruh H. sapiens ve druhé polovině 20. století. Odvozeno od konvence, rozšířené v 80. letech, uvažování o dvou poddruhech, H. s. neanderthalensis a H. s. sapiens, výslovné tvrzení, že „H. s. sapiens je jediný existující lidský poddruh “se objevuje na počátku 90. let.[55]
Od 2000s, zaniklý Homo sapiens idaltu (Bílý et al., 2003) získal široké uznání jako poddruh Homo sapiens, ale i v tomto případě existuje nesouhlasný názor, který tvrdí, že „lebky nemusí být natolik výrazné, aby vyžadovaly nový název poddruhu“.[56] H. s. neanderthalensis a H. s. rhodesiensis některé úřady nadále považují za samostatný druh, ale objev genetického důkazu o archaická příměs člověka s moderními lidmi znovu otevřel podrobnosti taxonomie archaických lidí.[57]
Homo erectus poddruh
Homo erectus od svého zavedení v roce 1892 byl rozdělen do mnoha poddruhů, z nichž mnohé byly dříve považovány za jednotlivé druhy Homo. Žádný z těchto poddruhů nemá mezi paleontology všeobecnou shodu.
- Homo erectus erectus (Java Man ) (70. léta)[58]
- Homo erectus yuanmouensis (Yuanmou Man ) (Li et al., 1977)
- Homo erectus lantianensis (Lantian Man ) (Woo Ju-Kang, 1964)
- Homo erectus nankinensis (Nanjing Man ) (1993)
- Homo erectus pekinensis (Peking Man ) (70. léta)[58]
- Homo erectus palaeojavanicus (Meganthropus ) (Tyler, 2001)
- Homo erectus soloensis (Solo Man ) (Oppenoorth, 1932)
- Homo erectus tautavelensis (Tautavel Man ) (de Lumley and de Lumley, 1971)
- Homo erectus georgicus (1991)
- Homo erectus bilzingslebenensis (Vlček, 2002)[59]
Viz také
Poznámky pod čarou
- ^ Potvrzeno H. habilis fosilie jsou datovány před 2,1 až 1,5 miliony let. Toto časové období se překrývá se vznikem Homo erectus.[22][23]
- ^ Homininy se znaky „proto-Homo“ mohly žít již před 2,8 miliony let, jak naznačuje fosilní čelistní kost klasifikovaná jako přechodná mezi Australopithecus a Homo objeveno v roce 2015.
- ^ Druh navržený v roce 2010 na základě fosilních pozůstatků tří jedinců datovaných před 1,9 až 0,6 miliony let. Stejné fosílie byly také klasifikovány jako H. habilis, H. ergaster nebo Australopithecus jinými antropology.
- ^ H. erectus se mohly objevit asi před 2 miliony let. Fosílie datované před 1,8 miliony let byly nalezeny jak v Africe, tak v jihovýchodní Asii a nejstarší fosilie s úzkým okrajem (před 1,85 až 1,77 miliony let) byly nalezeny na Kavkaze, takže není jasné, zda H. erectus se objevily v Africe a migrovaly do Eurasie, nebo pokud se naopak vyvinuly v Eurasii a migrovaly zpět do Afriky.
- ^ Nyní také součástí H. erectus jsou Peking Man (dříve Sinanthropus pekinensis) a Java Man (dříve Pithecanthropus erectus).
- ^ H. erectus je nyní seskupena do různých poddruhů, včetně Homo erectus erectus, Homo erectus yuanmouensis, Homo erectus lantianensis, Homo erectus nankinensis, Homo erectus pekinensis, Homo erectus palaeojavanicus, Homo erectus soloensis, Homo erectus tautavelensis, Homo erectus georgicus. Rozdíl od potomků, jako jsou Homo ergaster, Homo floresiensis, Homo předchůdce, Homo heidelbergensis a vskutku Homo sapiens není zcela jasné.
- ^ Typ fosílie je Mauer 1, datováno do ca. Před 0,6 miliony let. Přechod z H. heidelbergensis na H. neanderthalensis před 300 až 243 tisíci lety je konvenční a využívá skutečnosti, že v tomto období nejsou známy žádné fosilie. Příklady H. heidelbergensis jsou fosilie nalezené na Bilzingsleben (také klasifikováno jako Homo erectus bilzingslebensis).
- ^ Věk H. sapiens dlouho se předpokládalo, že se blíží 200 000 let, ale od roku 2017 existuje řada návrhů, které tuto dobu prodlužují až na 300 000 let. v roce 2017 byly fosilie nalezené v Jebel Irhoud (Maroko ) navrhuji, aby Homo sapiens možná se specializovali již před 315 000 lety.[31]Genetické důkazy byly předloženy pro věk zhruba 270 000 let.[32]
- ^ První lidé s „proto-neandertálskými vlastnostmi“ žili v Eurasii již před 0,6 až 0,35 miliony let (klasifikováno jako H. heidelbergensis, také nazývaný a chronospecies protože představuje spíše chronologické seskupení, než aby bylo založeno na jasných morfologických odlišnostech od obou H. erectus nebo H. neanderthalensis). V Evropě existuje fosilní propast mezi 300 a 243 kya a podle konvencí se fosilie mladší než 243 kya nazývají „neandertálci“.[34]
- ^ mladší než 450 kya, buď mezi 190–130 nebo mezi 70–10 kya[35]
- ^ prozatímní jména Homo sp. Altaj nebo Homo sapiens ssp. Denisova.
- ^ Bølling – oteplování Allerød doba
Reference
- ^ Stringer, Chris (12. června 2003). „Lidská evoluce: z Etiopie“. Příroda. 423 (6941): 693–695. Bibcode:2003 Natur.423..692S. doi:10.1038 / 423692a. PMID 12802315. S2CID 26693109.
- ^ „Herto lebky (Homo sapiens idaltu)“. talkorigins org. Citováno 7. června 2016.
- ^ Stringer, C. (2016). „Původ a vývoj Homo sapiens“. Filozofické transakce Královské společnosti v Londýně. Série B, Biologické vědy. 371 (1698): 20150237. doi:10.1098 / rstb.2015.0237. PMC 4920294. PMID 27298468.
- ^ Jared Diamond v Třetí šimpanz (1991) a Morris Goodman (2003)Hecht, Jeff (19. května 2003). „Šimpanzi jsou lidé, naznačuje to genová studie“. Nový vědec. Citováno 2011-12-08.
- ^ K. Wagner, Jennifer (2016). „Názory antropologů na rasu, předky a genetiku“. American Journal of Physical Anthropology. 162 (2): 318–327. doi:10.1002 / ajpa.23120. PMC 5299519. PMID 27874171.
- ^ „AAA Statement on Race“. Americká antropologická asociace.
- ^ J. E. Gray, „Nástin pokusu o rozmístění Mammalia do kmenů a rodin, se seznamem rodů zjevně vhodných pro každý kmen“, Annals of Philosophy, nová řada (1825), str. 337–344.
- ^ Cela-Conde, C. J .; Ayala, F. J. (2003). "Rody lidské linie". Sborník Národní akademie věd. 100 (13): 7684–7689. Bibcode:2003PNAS..100.7684C. doi:10.1073 / pnas.0832372100. PMC 164648. PMID 12794185.
- ^ Představeno pro Florisbadská lebka (objeveno v roce 1932, Homo florisbadensis nebo Homo helmei). Rod také navrhl pro řadu archaických lidských lebek nalezených v Jezero Eyasi Weinert (1938). Děravý, Journal of the East Africa Natural History Society (1942), p. 43.
- ^ Villmoare, B. (2015). „Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia“. Věda. 347 (6228): 1352–1355. Bibcode:2015Sci ... 347.1352V. doi:10.1126 / science.aaa1343. PMID 25739410.. Někteří paleoantropologové se domnívají H. habilis taxon jako neplatný, složený z fosilních exemplářů Australopithecus a Homo. Tattersall, I. & Schwartz, J.H., Zaniklý člověk, Westview Press, New York, 2001, s. 111.
- ^ De Heinzelin, J; Clark, JD; Bílá, T; Hart, W; Renne, P; Woldegabriel, G; Beyene, Y; Vrba, E (1999). „Prostředí a chování 2,5 milionu let starých Bouri hominidů“. Věda. 284 (5414): 625–9. Bibcode:1999Sci ... 284..625D. doi:10.1126 / science.284.5414.625. PMID 10213682.
- ^ Kaplan, Matt (8. srpna 2012). „Fosílie ukazují na velkou rodinu předků lidí“. Příroda. Citováno 8. srpna 2012.
- ^ Wood a Richmond; Richmond, BG (2000). „Vývoj člověka: taxonomie a paleobiologie“. Anatomy Journal. 197 (Pt 1): 19–60. doi:10.1046 / j.1469-7580.2000.19710019.x. PMC 1468107. PMID 10999270.
- ^ Reynolds, Sally C; Gallagher, Andrew (29.03.2012). African Genesis: Perspectives on Hominin Evolution. ISBN 9781107019959.
- ^ Brunet, M .; et al. (2002). „Nový hominid z horního miocénu v Čadu ve střední Africe“. Příroda. 418 (6894): 145–151. Bibcode:2002 Natur.418..145B. doi:10.1038 / nature00879. PMID 12110880. S2CID 1316969.Cela-Conde, C.J .; Ayala, F.J. (2003). "Rody lidské linie". PNAS. 100 (13): 7684–7689. Bibcode:2003PNAS..100.7684C. doi:10.1073 / pnas.0832372100. PMC 164648. PMID 12794185.Wood, B .; Lonergan, N. (2008). „Fosilní záznam homininů: taxony, stupně a subtypy“ (PDF). J. Anat. 212 (4): 354–376. doi:10.1111 / j.1469-7580.2008.00871.x. PMC 2409102. PMID 18380861.
- ^ Hazarika, Manji (16. – 30. Června 2007). "Homo erectus / ergaster a mimo Afriku: Nedávný vývoj v paleoantropologii a prehistorické archeologii " (PDF).
- ^ Klein, R. (1999). Kariéra člověka: Lidský biologický a kulturní původ. Chicago, IL: University of Chicago Press. ISBN 0226439631.
- ^ Antón, S.C. (2003). "Přírodní historie Homo erectus". Dopoledne. J. Phys. Anthropol. 122: 126–170. doi:10.1002 / ajpa.10399. PMID 14666536.
V 80. letech rostl počet H. erectus vzorky, zejména v Africe, vedly k poznání, že asijské H. erectus (H. erectus sensu stricto), kdysi považovaný za primitivní, byl ve skutečnosti odvozenější než jeho africké protějšky. Někteří tyto morfologické rozdíly interpretovali jako důkaz, že do nich může být zahrnut více než jeden druh H. erectus sensu lato (např. Stringer, 1984; Andrews, 1984; Tattersall, 1986; Wood, 1984, 1991a, b; Schwartz a Tattersall, 2000) ... Na rozdíl od evropské linie, podle mého názoru, taxonomické problémy kolem asijských vs. afrických H erectus jsou nepoddajnější. Tato otázka byla nejvíce ostře řešena pojmenováním H. ergaster na základě typové čelisti KNM-ER 992, ale také zahrnující částečnou kostru a izolované zuby KNM-ER 803 mezi ostatními pozůstatky Koobi Fora (Groves a Mazak, 1975 ). Nedávno byl tento specifický název používán u většiny raných afrických a gruzínských H. erectus jako uznání méně odvozené povahy těchto ostatků ve vztahu k podmínkám v asijském H. erectus (viz Wood, 1991a, s. 268; Gabunia et al. , 2000a). Alespoň části paratypu H. ergaster (např. KNM-ER 1805) nejsou zahrnuty ve většině současných koncepcí daného taxonu. The H. ergaster otázka zůstává skvěle nevyřešena (např. Stringer, 1984; Tattersall, 1986; Wood, 1991a, 1994; Rightmire, 1998b; Gabunia et al., 2000a; Schwartz a Tattersall, 2000), v nemalé míře, protože původní diagnóza neposkytovala žádné srovnání s asijskými fosilními záznamy.
- ^ „Lebka naznačuje, že tři rané lidské druhy byly jeden“. Novinky a komentáře. Příroda.
- ^ Lordkipanidze, David; Ponce de Leòn, Marcia S .; Margvelashvili, Ann; Rak, Yoel; Rightmire, G. Philip; Vekua, Abesalom; Zollikofer, Christoph P. E. (18. října 2013). "Kompletní lebka z Dmanisi v Gruzii a evoluční biologie raného Homo". Věda. 342 (6156): 326–331. Bibcode:2013Sci ... 342..326L. doi:10.1126 / science.1238484. PMID 24136960. S2CID 20435482.
- ^ Switek, Brian (17. října 2013). „Krásná debata o lebce o lidských dějinách“. národní geografie. Citováno 22. září 2014.
- ^ Schrenk F, Kullmer O, Bromage T (2007). „Nejdříve domnělý Homo Fosílie ". V Henke W, Tattersall I. (eds.). Příručka paleoantropologie. 1. Ve spolupráci s Thorolfem Hardtem. Berlín, Heidelberg: Springer. 1611–1631. doi:10.1007/978-3-540-33761-4_52. ISBN 978-3-540-32474-4.
- ^ DiMaggio EN, Campisano CJ, Rowan J, Dupont-Nivet G, Deino AL, Bibi F a kol. (Březen 2015). "Paleoanthropology. Pozdní pliocénní fosiliferous sedimentární záznam a environmentální kontext raného Homo z Afar, Etiopie". Věda. 347 (6228): 1355–9. Bibcode:2015Sci ... 347.1355D. doi:10.1126 / science.aaa1415. PMID 25739409. S2CID 43455561.
- ^ Curnoe D (červen 2010). „Recenze raného Homo v jižní Africe se zaměřením na lebeční, mandibulární a zubní pozůstatky s popisem nového druhu (Homo gautengensis sp. Nov.)“. Homo. 61 (3): 151–77. doi:10.1016 / j.jchb.2010.04.002. PMID 20466364.
- ^ Haviland WA, Walrath D, Prins HE, McBride B (2007). Evolution and Prehistory: The Human Challenge (8. vydání). Belmont, CA: Thomson Wadsworth. p. 162. ISBN 978-0-495-38190-7.
- ^ Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T a kol. (Červen 2011). „Nejstarší lidské okupace v Dmanisi (gruzínský Kavkaz) se datovaly 1,85–1,78 Ma“. Sborník Národní akademie věd Spojených států amerických. 108 (26): 10432–6. Bibcode:2011PNAS..10810432F. doi:10.1073 / pnas.1106638108. PMC 3127884. PMID 21646521.
- ^ Indriati E, Swisher CC, Lepre C, Quinn RL, Suriyanto RA, Hascaryo AT a kol. (2011). „Věk 20metrové terasy řeky Solo, Jáva, Indonésie a přežití Homo erectus v Asii“. PLOS ONE. 6 (6): e21562. Bibcode:2011PLoSO ... 621562I. doi:10.1371 / journal.pone.0021562. PMC 3126814. PMID 21738710.
- ^ Hazarika M (2007). "Homo erectus / ergaster a mimo Afriku: Nedávný vývoj v paleoantropologii a prehistorické archeologii " (PDF). EKniha letní školy EAA. 1. Evropská antropologická asociace. str. 35–41.
Intenzivní kurz biologické antrpologie, 1. letní škola Evropské antropologické asociace, 16. – 30. Června 2007, Praha, Česká republika
- ^ Muttoni G, Scardia G, Kent DV, Swisher CC, Manzi G (2009). "Pleistocene magnetochronology of early hominin sites at Ceprano and Fontana Ranuccio, Italy". Dopisy o Zemi a planetách. 286 (1–2): 255–268. Bibcode:2009E & PSL.286..255M. doi:10.1016 / j.epsl.2009.06.032.
- ^ Dirks PH, Roberts EM, Hilbert-Wolf H, Kramers JD, Hawks J, Dosseto A, et al. (Květen 2017). „Homo naledi a související sedimenty v jeskyni Rising Star, Jihoafrická republika“. eLife. 6: e24231. doi:10,7554 / eLife.24231. PMC 5423772. PMID 28483040.
- ^ Callaway, Ewan (7. června 2017). „Nejstarší fosilní tvrzení Homo sapiens přepisuje historii našeho druhu“. Příroda. doi:10.1038 / příroda.2017.22114. Citováno 11. června 2017.
- ^ Posth C, Wißing C, Kitagawa K, Pagani L, van Holstein L, Racimo F a kol. (Červenec 2017). „Hluboce divergentní archaický mitochondriální genom poskytuje nižší časovou hranici pro tok afrických genů do neandertálců“. Příroda komunikace. 8: 16046. Bibcode:2017NatCo ... 816046P. doi:10.1038 / ncomms16046. PMC 5500885. PMID 28675384.
- ^ Bischoff JL, Shamp DD, Aramburu A a kol. (Březen 2003). „The Sima de los Huesos Hominids Date to Beyond U / Th Equilibrium (> 350 kyr) and Možná na 400–500 kyr: New Radiometric Dates“. Journal of Archaeological Science. 30 (3): 275–280. doi:10.1006 / jasc.2002.0834. ISSN 0305-4403.
- ^ Dean D, Hublin JJ, Holloway R, Ziegler R (květen 1998). „O fylogenetické poloze předeandertálského vzorku z německého Reilingenu.“ Journal of Human Evolution. 34 (5): 485–508. doi:10.1006 / jhev.1998.0214. PMID 9614635.
- ^ Chang CH, Kaifu Y, Takai M, Kono RT, Grün R, Matsu'ura S a kol. (Leden 2015). „První archaický Homo z Tchaj-wanu“. Příroda komunikace. 6: 6037. Bibcode:2015NatCo ... 6,6037C. doi:10.1038 / ncomms7037. PMC 4316746. PMID 25625212.
- ^ Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E a kol. (Duben 2019). "Nový druh Homo z pozdního pleistocénu na Filipínách". Příroda. 568 (7751): 181–186. Bibcode:2019Natur.568..181D. doi:10.1038 / s41586-019-1067-9. PMID 30971845. S2CID 106411053.
- ^ Zimmer C. (10. dubna 2019). „V této filipínské jeskyni kdysi žil nový lidský druh - archeologové na Luzonském ostrově objevili kosti vzdáleného příbuzného druhu Homo luzonensis, čímž dále rozšířili lidský rodokmen“. The New York Times. Citováno 10. dubna 2019.
- ^ Curnoe D, Xueping J, Herries AI, Kanning B, Taçon PS, Zhende B a kol. (2012). „Lidské ostatky z přechodu pleistocén-holocén v jihozápadní Číně naznačují pro východní Asijce složitou evoluční historii“. PLOS ONE. 7 (3): e31918. Bibcode:2012PLoSO ... 731918C. doi:10.1371 / journal.pone.0031918. PMC 3303470. PMID 22431968.
- ^ „pokud vím, neexistuje žádný typový materiál Homo sapiens. Abychom byli k Linné spravedliví, nezdálo se, že by se praxe odkládání vzorků typu odvíjela až o sto let později. “Bob Ralph,„ V souladu s typem Nový vědec Č. 1548 (19. února 1987), p. 59.
- ^ „Glosář ICZN“. Mezinárodní kodex zoologické nomenklatury. 4. vydání, článek 46.1: „Prohlášení o zásadě koordinace vztahující se na názvy skupin druhů. Název stanovený pro taxon na kterékoli pozici ve skupině druhů je považován za název, který byl současně vytvořen stejným autorem pro taxon na jiné pořadí ve skupině; oba nominální taxony mají stejný typ nesoucí jméno, ať už byl tento typ opraven původně nebo později. “Homo sapiens sapiens je zřídka používán před čtyřicátými léty. V roce 1946 John Wendell Bailey výslovně připisuje jméno Linné (1758): „Linné. Syst. Nat. Ed. 10, sv. 1. str. 20, 21, 22, uvádí pět ras člověka, viz: Homo sapiens sapiens (bílý - bělošský) [...] ", Toto je chybné přiřazení, ale H. s. sapiens Od té doby se často připisuje Linné. Ve skutečnosti Linné, Syst. Nat. vyd. 10 sv. 1. p. 21 nemá Homo sapiens sapiensmísto toho se nazývá „bílá“ nebo „bělošská“ rasa Homo sapiens Europaeus. To je výslovně uvedeno v Bulletin der Schweizerische Gesellschaft für Anthropologie und Ethnologie Svazek 21 (1944), str. 18 (argumentovat proti H. s. sapiens ale proti "H. s. albus L.„navrhli von Eickstedt a Peters):“ die europide Rassengruppe, als Subspecies aufgefasst, [würde] Homo sapiens eurpoaeus L. heissen "(" rasová skupina Europid, považovaná za poddruh, by byla pojmenována H. s. europeaeus L."). Viz také: John R. Baker, Závod, Oxford University Press (1974), 205.
- ^ Stearn, W. T. 1959. „Pozadí Linnéových příspěvků k nomenklatuře a metodám systematické biologie“, Systematic Zoology 8 (1): 4-22, s. 1. 4
- ^ „Je jisté, že Linné byl přítomen, když psal tento popis, a že se považoval za člena Homo sapiens. Že není součástí žádné z jeho podskupin, je zřejmé z popisů, zejména že určitě není součástí Homo sapiens europaeus, protože tento poddruh je popsán jako „Pilis flavescentibus, prolixis. Oculis caeruleis“, zatímco Linnaeus má hnědé vlasy a oči (Tullberg, 1907). Je proto zahrnut do typové řady Homo sapiens sapiens (článek 72.4.1.1). Žádná jednotlivá osoba však nebyla uznána za typ až do roku 1959, kdy profesor William Stearn v komentáři v příspěvku k Linnéovým příspěvkům k nomenklatuře a systematice napsal, že „sám Linné musí být typem svého Homo sapiens“. To stačilo k označení Linné jako lektotypu (článek 74.5), jediného exempláře nesoucího typ pro druh Homo sapiens a jeho poddruh Homo sapiens sapiens. “David Notton a Chris Stringer, „Kdo je typ Homo sapiens?“, Mezinárodní komise pro zoologickou nomenklaturu (2011).
- ^ Linné, Carl von (1758). Systema naturæ. Regnum animale (10. vyd.). 18 s.
- ^ Viz např. John Wendell Bailey, Savci ve Virginii (1946), str. 356 .; Journal of Mammalogy 26-27 (1945), str. 359.; J. Desmond Clark (ed.), Cambridge historie Afriky, Cambridge University Press (1982), p. 141 (s odkazy).
- ^ Annals of Philosophy 11, Londýn (1826), p. 71
- ^ Frederick S. Szalay, Eric Delson, Evoluční historie primátů (2013), 508
- ^ Pääbo, Svante (2014). Neandertálský muž: Hledání ztracených genomů. New York: Základní knihy. p. 237.
- ^ Groves, C. P. (2005). Wilson, D. E.; Reeder, D. M. (eds.). Savčí druhy světa: taxonomický a zeměpisný odkaz (3. vyd.). Baltimore: Johns Hopkins University Press. ISBN 0-801-88221-4. OCLC 62265494.
- ^ T. Harrison in: William H. Kimbel, Lawrence B. Martin (eds.), Druhy, pojmy druhů a vývoj primátů (2013), 361.
- ^ M. R. Drennan, „Australoidská lebka z Cape Flats“, The Journal of the Royal Anthropological Institute of Great Britain and Ireland Sv. 59 (červenec - prosinec, 1929), 417-427.
- ^ mimo jiné navrhovaná jména pro fosilie později zahrnutá pod neanderthalensisviz: Eric Delson, Ian Tattersall, John Van Couvering, Alison S. Brooks, Encyclopedia of Human Evolution and Prehistory: Second Edition, Routledge (2004).
- ^ GP Rightmire (3. června 1983). „Jezero Ndutu lebka a brzy Homo sapiens v Africe". Dopoledne. J. Phys. Anthropol. 61 (2): 245–54. doi:10.1002 / ajpa.1330610214. PMID 6410925.
- ^ Anglický překlad (1972–1975): Grzimekova encyklopedie o životě zvířat, Svazek 11, str. 55.
- ^ John R. Baker, Závod, Oxford University Press (1974).
- ^ „Jsme jediný přežívající poddruh Homo sapiens"Michio Kitahara, Tragédie evoluce: lidské zvíře čelí moderní společnosti (1991), str. xi.
- ^ Chris Stringer (12. června 2003). „Vývoj člověka: z Etiopie“. Příroda.
- ^ Hublin, J. J. (2009). „Původ neandertálců“. Sborník Národní akademie věd. 106 (38): 16022–7. Bibcode:2009PNAS..10616022H. doi:10.1073 / pnas.0904119106. JSTOR 40485013. PMC 2752594. PMID 19805257.Harvati, K .; Frost, S.R .; McNulty, K.P. (2004). "Přehodnocena neandertálská taxonomie: důsledky 3D modelů primátů pro vnitrodruhové a mezidruhové rozdíly". Proc. Natl. Acad. Sci. USA. 101 (5): 1147–52. Bibcode:2004PNAS..101.1147H. doi:10.1073 / pnas.0308085100. PMC 337021. PMID 14745010."Homo neanderthalensis King, 1864 ". Wiley-Blackwell Encyclopedia of Human Evolution. Chichester, West Sussex: Wiley-Blackwell. 2013. s. 328–331.
- ^ A b V 70. letech se vyvinula tendence k jávské odrůdě H. erectus jako poddruh, Homo erectus erectus, přičemž čínská odrůda je označována jako Homo erectus pekinensis. Viz: Sartono, S. Důsledky vyplývající z Pithecanthropus VIII In: Paleoantropologie: morfologie a paleoekologie. Russell H. Tuttle (ed.), Str. 328.
- ^ Emanuel Vlček: Der fosilní Mensch von Bilzingsleben (= Bilzingsleben. Bd. 6 = Beiträge zur Ur- und Frühgeschichte Mitteleuropas 35). Beier & Beran, Langenweißbach 2002.
| Taxonomie (Homininy ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Předci |
| ||||||||||||||||||||||||||
| Modely |
| ||||||||||||||||||||||||||
| Časové osy | |||||||||||||||||||||||||||
| Ostatní |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||