Embryoidní tělo - Embryoid body
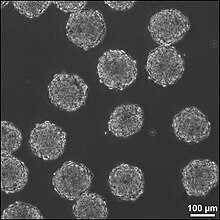

Embryoidní těla (EB) jsou trojrozměrné agregáty pluripotentu kmenové buňky.
EB jsou diferenciací lidských embryonálních kmenových buněk na embryoidní těla, která ohrožuje tři embryonální zárodečné vrstvy.
Pozadí
Mezi pluripotentní typy buněk, které obsahují embryoidní těla, patří embryonální kmenové buňky (ESC) odvozené z blastocyst stadium embryí z myší (mESC),[1][2] primát,[3] a člověk (hESC)[4] Zdroje. Navíc mohou být EB vytvořeny z embryonálních kmenových buněk odvozených pomocí alternativních technik, včetně somatických buněk jaderný přenos[5][6][7] nebo přeprogramování somatických buněk na výtěžek indukované pluripotentní kmenové buňky (iPS).[8][9][10][11] Podobně jako ESC kultivované v monovrstvě formáty podléhají ESC v embryoidních tělech diferenciaci a specifikaci buněk podél těchto tří rodové linie - endoderm, ektoderm, a mesoderm - které zahrnují všechny somatický typy buněk.[12][13]
Na rozdíl od monovrstevných kultur však sféroidní struktury, které se tvoří, když se agregují ESC, umožňují neadherentní kulturu EB v suspenzi, čímž jsou kultury EB inherentně škálovatelné, což je užitečné pro přístupy k biologickému zpracování, čímž lze produkovat velké výtěžky buněk potenciální klinické aplikace.[14] Navíc, i když EB do značné míry vykazují heterogenní vzorce diferencovaných typů buněk, jsou ESC schopné reagovat na podobné podněty, které řídí embryonální vývoj.[15] Proto je trojrozměrná struktura, včetně vytvoření komplexu buněčné adheze a parakrin signalizace v mikroprostředí EB,[16] umožňuje rozlišení a morfogeneze který poskytuje mikrotkánky podobné nativním tkáňovým strukturám. Takové mikrotlaky slibují přímo[15] nebo nepřímo[17][18] opravovat poškozené nebo nemocné tkáně v aplikacích regenerativní medicíny, jakož i pro testování in vitro ve farmaceutickém průmyslu a jako model embryonálního vývoje.
Formace
EB jsou tvořeny homofilní vazba Ca2 + závislé adhezní molekuly E-kadherin, což je vysoce vyjádřeno na nediferencovaných HSR.[19][20][21] Pokud jsou kultivovány jako jednotlivé buňky za nepřítomnosti anti-diferenciačních faktorů, ESC se spontánně agregují za vzniku EB.[19][22][23][24] Takové spontánní tvorby se často dosahuje v objemových suspenzních kulturách, kdy je miska potažena nelepivými materiály, jako je např agar nebo hydrofilní polymery, aby se podpořila preferenční adheze mezi jednotlivými buňkami spíše než ke kultivačnímu substrátu. Protože hESC podléhá apoptóze, je-li pěstována jako jednotlivé buňky, tvorba EB často vyžaduje použití inhibitorů kináza spojená s rho (ROCK) dráha, včetně malých molekul Y-27632[25] a 2,4 disubstituovaný thiazol (Thiazovivin / Tzv).[26] Alternativně, aby se zabránilo disociaci na jednotlivé buňky, mohou být EB vytvořeny z hESC pomocí manuální separace adherentních kolonií (nebo oblastí kolonií) a následně kultivovány v suspenzi. Tvorba EB v suspenzi je přístupná tvorbě velkého množství EB, ale poskytuje malou kontrolu nad velikostí výsledných agregátů, což často vede k velkým EB nepravidelného tvaru. Jako alternativu hydrodynamický síly předávané na platformách smíšené kultury zvyšují homogenitu velikostí EB, když jsou ESC naočkovány do hromadných suspenzí.[27]
Tvorba EB může být také přesněji řízena naočkováním známých buněčných hustot v jednotlivých kapkách (10-20 ul) suspendovaných z víčka Petriho misky, známých jako visící kapky.[21] I když tato metoda umožňuje kontrolu velikosti EB změnou počtu buněk na kapku, tvorba visících kapek je náročná na práci a není snadno přizpůsobitelná škálovatelným kulturám. Média navíc nelze snadno vyměnit v tradičním formátu zavěšených kapek, což vyžaduje přenos zavěšených kapek do objemových suspenzních kultur po 2–3 dnech tvorby, přičemž jednotlivé EB mají tendenci aglomerovat. Nedávno byly vyvinuty nové technologie umožňující výměnu médií v upraveném formátu visícího přetažení.[28] Kromě toho byly vyvinuty technologie pro fyzickou separaci buněk nucenou agregací ESC v jednotlivých jamkách nebo omezenou na adhezivní substráty,[29][30][31][32] což umožňuje zvýšenou propustnost, kontrolovanou tvorbu EB. Nakonec mohou metody použité pro tvorbu EB ovlivnit heterogenitu populací EB, pokud jde o kinetiku agregace, velikost a výtěžek EB, stejně jako trajektorie diferenciace.[31][33][34]
Diferenciace v rámci EB
V kontextu ESC diferenciace protokoly, tvorba EB se často používá jako metoda pro zahájení spontánní diferenciace směrem k těmto třem rodové linie. EB diferenciace začíná specifikací vnějších buněk směrem k primitivnímu fenotypu endodermu.[35][36] Buňky na vnější straně se poté usadí extracelulární matrix (ECM), obsahující kolagen IV a laminin,[37][38] podobné složení a struktuře bazální membrána. V reakci na ECM depozici, EB často tvoří cystickou dutinu, přičemž buňky v kontaktu se bazální membránou zůstávají životaschopné a buňky uvnitř procházejí apoptózou, což vede k dutině naplněné tekutinou obklopené buňkami.[39][40][41] Následná diferenciace pokračuje za vzniku derivátů tří zárodečných linií. Při absenci doplňků je „výchozí“ diferenciace ESC do značné míry k ektodermu a následná neurální linie.[42] Alternativní mediální kompozice, včetně použití fetální hovězí sérum stejně jako definované přísady růstových faktorů byly vyvinuty na podporu diferenciace k mezoderm a endoderm linie.[43][44][45]
V důsledku trojrozměrné struktury EB dochází během diferenciace EB ke složité morfogenezi, včetně vzhledu populací buněk podobných epiteliálním i mezenchymálním, stejně jako vzhledu markerů spojených s epiteliálně-mezenchymální přechod (EMT).[46][47] Induktivní účinky vyplývající ze signalizace mezi buněčnými populacemi v EB navíc vedou k prostorově a časově definovaným změnám, které podporují komplexní morfogeneze.[48] V EB jsou často vystaveny tkáňové struktury, včetně vzhledu krevních ostrovů připomínajících časné struktury krevních cév ve vyvíjejícím se embryu, stejně jako vzorování neurit rozšíření (svědčící o organizaci neuronů) a spontánní kontraktilní aktivita (svědčící o kardiomyocyt diferenciace), když jsou EB naneseny na adhezivní podklady, jako např želatina.[13] Více nedávno, komplexní struktury, včetně optických miskovitých struktur byly vytvořeny in vitro jako výsledek EB diferenciace.[49]
Paralely s embryonálním vývojem
Velká část výzkumu zaměřeného na diferenciaci a morfogenezi embryonálních kmenových buněk je odvozena ze studií vývojové biologie a embryogeneze savců.[15] Například bezprostředně po blastocyst ve stadiu vývoje (z něhož jsou odvozeny ESC) embryo prochází gastrulace, přičemž specifikace buňky vnitřní buněčná hmota vede k tvorbě viscerálního endodermu a epiblast.[50] Jako přední a zadní osa je vytvořeno, embryo vyvíjí přechodnou strukturu známou jako primitivní pruh.[51] Hodně z prostorového vzorování, ke kterému dochází během tvorby a migrace primitivního pruhu, je výsledkem sekrece agonisté a antagonisté různými populacemi buněk, včetně růstových faktorů z Wnt a transformující růstový faktor β (TGFβ) rodiny (Lefty 1, Nodal), stejně jako represory stejných molekul (Dkk-1, Sfrp1, Sfrp5).[52][53][54] Vzhledem k podobnostem mezi embryogenezí a ESC diferenciací je mnoho stejných růstových faktorů ústředním bodem pro přístupy řízené diferenciace.
Kromě toho vylepšení kultury EB vyústilo ve vývoj embryonální organoidy (gastruloidy) které vykazují pozoruhodné paralely s embryonálním vývojem[55][56][57][58][59] například rozbití symetrie, lokalizované brachyury výraz, vznik embryonální osy (předozadní, dorzoventrální a zleva doprava) a pohyby podobné gastrulaci.[55][56][57]
Výzvy směřující k diferenciaci
Na rozdíl od diferenciace ESC v jednovrstvých kulturách, kdy lze přesně a homogenně kontrolovat přidání rozpustných morfogenů a extracelulárního mikroprostředí, představuje trojrozměrná struktura EB výzvy pro cílenou diferenciaci.[16][60] Například viscerální populace endodermu, která tvoří vnějšek EB, vytváří vnější „skořápku“ skládající se z těsně spojených epiteliální - jako buňky, stejně jako hustá ECM.[61][62] Kvůli takovým fyzickým omezením v kombinaci s velikostí EB doprava v EB dochází k omezením, které vytvářejí přechody morfogenů, metabolitů a živin.[60] Odhaduje se, že transport kyslíku je omezen v buněčných agregátech větších než přibližně 300 um v průměru;[63] vývoj takových gradientů je však také ovlivněn velikostí molekul a rychlostí absorpce buněk. Proto dodávka morfogenů do EB vede ke zvýšení heterogenity a snížení účinnosti diferencovaných buněčných populací ve srovnání s monovrstvovými kulturami. Jedním ze způsobů řešení transportních omezení v rámci EB byl polymerní dodávání morfogenů ze struktury EB.[61][64][65] Navíc mohou být EB kultivovány jako jednotlivé mikrotkaničky a následně sestaveny do větších struktur pro aplikace tkáňového inženýrství.[66] Ačkoli složitost vyplývající z trojrozměrných adhezí a signalizace může rekapitulovat přirozenější tkáňové struktury,[67][68] také vytváří výzvy pro pochopení relativního příspěvku mechanických, chemických a fyzikálních signálů k výsledným buněčným fenotypům a morfogenezi.
Viz také
Reference
- ^ Martin, G. R. (1981). „Izolace pluripotentní buněčné linie z časných myších embryí kultivovaných v médiu upraveném kmenovými buňkami teratokarcinomu“. Sborník Národní akademie věd Spojených států amerických. 78 (12): 7634–7638. doi:10.1073 / pnas.78.12.7634. PMC 349323. PMID 6950406.
- ^ Evans, M. J .; Kaufman, M. H. (1981). "Založení pluripotenciálních buněk z myších embryí v kultuře". Příroda. 292 (5819): 154–156. doi:10.1038 / 292154a0. PMID 7242681.
- ^ Thomson, J. A .; Kalishman, J .; Golos, T. G .; Durning, M .; Harris, C. P .; Becker, R. A .; Hearn, J. P. (1995). "Izolace linie embryonálních kmenových buněk primátů". Sborník Národní akademie věd Spojených států amerických. 92 (17): 7844–7848. doi:10.1073 / pnas.92.17.7844. PMC 41242. PMID 7544005.
- ^ Thomson, J. A .; Itskovitz-Eldor, J .; Shapiro, S. S .; Waknitz, M. A .; Swiergiel, J. J .; Marshall, V. S .; Jones, J. M. (1998). „Embryonální linie kmenových buněk odvozené z lidských blastocyst“. Věda. 282 (5391): 1145–1147. doi:10.1126 / science.282.5391.1145. PMID 9804556.
- ^ Briggs, R .; King, T. J. (1952). „Transplantace živých jader z buněk blastuly do vajíček žabích vajec“. Sborník Národní akademie věd Spojených států amerických. 38 (5): 455–463. doi:10.1073 / pnas.38.5.455. PMC 1063586. PMID 16589125.
- ^ Wilmut, I.; Schnieke, A.E .; McWhir, J .; Kind, A. J .; Campbell, K. H. S. (1997). "Životaschopní potomci pocházející z buněk plodu a dospělých savců". Příroda. 385 (6619): 810–813. doi:10.1038 / 385810a0. PMID 9039911.
- ^ Munsie, M. J .; Michalska, A.E .; O'Brien, C. M .; Trounson, A. O .; Pera, M. F .; Mountford, P. S. (2000). "Izolace pluripotentních embryonálních kmenových buněk z přeprogramovaných jader somatických buněk dospělých myší". Aktuální biologie. 10 (16): 989–992. doi:10.1016 / s0960-9822 (00) 00648-5. PMID 10985386.
- ^ Takahashi, K .; Yamanaka, S. (2006). "Indukce pluripotentních kmenových buněk z myších embryonálních a dospělých fibroblastových kultur definovanými faktory". Buňka. 126 (4): 663–76. doi:10.1016 / j.cell.2006.07.024. hdl:2433/159777. PMID 16904174.
- ^ Takahashi, K .; Tanabe, K .; Ohnuki, M .; Narita, M .; Ichisaka, T .; Tomoda, K .; Yamanaka, S. (2007). "Indukce pluripotentních kmenových buněk z dospělých lidských fibroblastů definovanými faktory". Buňka. 131 (5): 861–872. doi:10.1016 / j.cell.2007.11.019. hdl:2433/49782. PMID 18035408.
- ^ Yu, J .; Vodyanik, M. A .; Smuga-Otto, K .; Antosiewicz-Bourget, J .; Frane, J. L .; Tian, S .; Nie, J .; Jonsdottir, G. A .; Ruotti, V .; Stewart, R .; Slukvin, I. I .; Thomson, J. A. (2007). "Indukované pluripotentní linie kmenových buněk odvozené z lidských somatických buněk". Věda. 318 (5858): 1917–1920. doi:10.1126 / science.1151526. PMID 18029452.
- ^ Park, I. H .; Arora, N .; Huo, H .; Maherali, N .; Ahfeldt, T .; Shimamura, A .; Lensch, M. W .; Cowan, C .; Hochedlinger, K .; Daley, G. Q. (2008). „Pluripotentní kmenové buňky vyvolané chorobou“. Buňka. 134 (5): 877–886. doi:10.1016 / j.cell.2008.07.041. PMC 2633781. PMID 18691744.
- ^ Itskovitz-Eldor, J .; Schuldiner, M .; Karsenti, D .; Eden, A .; Yanuka, O .; Amit, M .; Soreq, H .; Benvenisty, N. (2000). „Diferenciace lidských embryonálních kmenových buněk na embryoidní těla ohrožující tři embryonální zárodečné vrstvy“. Molekulární medicína (Cambridge, Massachusetts). 6 (2): 88–95. PMC 1949933. PMID 10859025.
- ^ A b Doetschman, T. C .; Eistetter, H .; Katz, M .; Schmidt, W .; Kemler, R. (1985). „In vitro vývoj linií embryonálních kmenových buněk odvozených z blastocyst: Tvorba viscerálního žloutkového vaku, krevních ostrovů a myokardu“. Journal of Embryology and Experimental Morfhology. 87: 27–45. PMID 3897439.
- ^ Dang, S. M .; Gerecht-Nir, S .; Chen, J .; Itskovitz-Eldor, J .; Zandstra, P. W. (2004). "Řízená, škálovatelná kultura diferenciace embryonálních kmenových buněk". Kmenové buňky. 22 (3): 275–282. doi:10.1634 / kmenové články.22-3-275. PMID 15153605.
- ^ A b C Murry, C.E .; Keller, G. (2008). „Diferenciace embryonálních kmenových buněk na klinicky relevantní populace: Poučení z embryonálního vývoje“. Buňka. 132 (4): 661–680. doi:10.1016 / j.cell.2008.02.008. PMID 18295582.
- ^ A b Bratt-Leal, A. S. M .; Carpenedo, R. L .; McDevitt, T. C. (2009). „Inženýrství mikroprostředí embryoidního těla k přímé diferenciaci embryonálních kmenových buněk“. Pokrok v biotechnologii. 25 (1): 43–51. doi:10,1002 / btpr.139. PMC 2693014. PMID 19198003.
- ^ Nair, R .; Shukla, S .; McDevitt, T. C. (2008). "Acelulární matice odvozené z diferenciace embryonálních kmenových buněk". Journal of Biomedical Materials Research Part A. 87A (4): 1075–1085. doi:10,1002 / jbm.a.31851. PMID 18260134.
- ^ Baraniak, P. R .; McDevitt, T. C. (2010). „Parakrinní působení kmenových buněk a regenerace tkání“. Regenerativní medicína. 5 (1): 121–143. doi:10,2217 / rme.09,74. PMC 2833273. PMID 20017699.
- ^ A b Kurosawa, H. (2007). "Metody vyvolání tvorby embryoidního těla: In vitro diferenciační systém embryonálních kmenových buněk". Journal of Bioscience and Bioengineering. 103 (5): 389–398. doi:10,1263 / jbb.103,389. PMID 17609152.
- ^ Larue, L .; Antos, C .; Butz, S .; Huber, O .; Delmas, V .; Dominis, M .; Kemler, R. (1996). "Role kadherinů při tvorbě tkáně". Rozvoj. 122 (10): 3185–3194. PMID 8898231.
- ^ A b Yoon, B. S .; Yoo, S. J .; Lee, J.E .; Ty, S .; Lee, H. T .; Yoon, H. S. (2006). "Zvýšená diferenciace lidských embryonálních kmenových buněk na kardiomyocyty kombinací zavěšené kapkové kultury a ošetření 5-azacytidinem". Diferenciace. 74 (4): 149–159. doi:10.1111 / j.1432-0436.2006.00063.x. PMID 16683985.
- ^ Park, J. H .; Kim, S.J .; E. J .; Moon, S. Y .; Roh, S. I .; Kim, C. G .; Yoon, H. S. (2003). „Vznik a údržba lidských embryonálních kmenových buněk na STO, trvale rostoucí buněčné linii“. Biologie reprodukce. 69 (6): 2007–2014. doi:10.1095 / biolreprod.103.017467. PMID 12930726.
- ^ Williams, R.L .; Hilton, D. J .; Pease, S .; Willson, T. A .; Stewart, C. L .; Gearing, D. P .; Wagner, E. F .; Metcalf, D .; Nicola, N. A .; Gough, N. M. (1988). „Faktor inhibující myeloidní leukémii udržuje vývojový potenciál embryonálních kmenových buněk“. Příroda. 336 (6200): 684–687. doi:10.1038 / 336684a0. PMID 3143916.
- ^ Ludwig, T. E.; Levenstein, M. E.; Jones, J. M .; Berggren, W. T .; Mitchen, E. R .; Frane, J. L .; Crandall, L. J .; Daigh, C. A .; Conard, K. R .; Piekarczyk, M. S .; Llanas, R. A .; Thomson, J. A. (2006). "Odvození lidských embryonálních kmenových buněk za definovaných podmínek". Přírodní biotechnologie. 24 (2): 185–187. doi:10.1038 / nbt1177. PMID 16388305.
- ^ Watanabe, K .; Ueno, M .; Kamiya, D .; Nishiyama, A .; Matsumura, M .; Wataya, T .; Takahashi, J. B .; Nishikawa, S .; Nishikawa, S. I .; Muguruma, K .; Sasai, Y. (2007). "ROCK inhibitor umožňuje přežití disociovaných lidských embryonálních kmenových buněk". Přírodní biotechnologie. 25 (6): 681–686. doi:10.1038 / nbt1310. PMID 17529971.
- ^ Xu, Y .; Zhu, X .; Hahm, H. S .; Wei, W .; Hao, E .; Hayek, A .; Ding, S. (2010). „Odhalení základního regulačního mechanismu signalizace pro přežití pluripotentních kmenových buněk a sebeobnovu malými molekulami“. Sborník Národní akademie věd. 107 (18): 8129–8134. doi:10.1073 / pnas.1002024107. PMC 2889586. PMID 20406903.
- ^ Carpenedo, R. L .; Sargent, C. Y .; McDevitt, T. C. (2007). „Kultura rotačního zavěšení zvyšuje účinnost, výtěžek a homogenitu diferenciace embryonálního těla“. Kmenové buňky. 25 (9): 2224–2234. doi:10.1634 / kmenové články 2006-0523. PMID 17585171.
- ^ Tung, Y. C .; Hsiao, A. Y .; Allen, S. G .; Torisawa, Y. S .; Ho, M .; Takayama, S. (2011). „Vysoce výkonná 3D sféroidní kultura a testování na drogy pomocí závěsného pole 384“. Analytik. 136 (3): 473–478. doi:10.1039 / c0an00609b. PMC 7454010. PMID 20967331. S2CID 35415772.
- ^ Park, J .; Cho, C.H .; Parashurama, N .; Li, Y .; Berthiaume, F. O .; Toner, M .; Tilles, A. W .; Yarmush, M. L. (2007). „Modulace diferenciace embryonálních kmenových buněk na základě mikrofabrikace“. Laboratoř na čipu. 7 (8): 1018–1028. doi:10.1039 / b704739h. PMID 17653344.
- ^ Mohr, J. C .; De Pablo, J. J .; Paleček, S. P. (2006). "3-D mikrojamková kultura lidských embryonálních kmenových buněk". Biomateriály. 27 (36): 6032–6042. doi:10.1016 / j.biomaterials.2006.07.012. PMID 16884768.
- ^ A b Hwang, Y. -S .; Chung, B.G .; Ortmann, D .; Hattori, N .; Moeller, H.C .; Khademhosseini, A. (2009). „Kontrola velikosti těla embrya zprostředkovaná mikrojamkami reguluje osud embryonálních kmenových buněk prostřednictvím diferenciální exprese WNT5a a WNT11“. Sborník Národní akademie věd. 106 (40): 16978–16983. doi:10.1073 / pnas.0905550106. PMC 2761314. PMID 19805103.
- ^ Ungrin, M. D .; Joshi, C .; Nica, A .; Bauwens, C.L .; Zandstra, P. W. (2008). Callaerts, Patrick (vyd.). „Reprodukovatelná tvorba ultravysokého výkonu mnohobuněčné organizace z agregátů lidských embryonálních kmenových buněk odvozených od jedné buňky“. PLOS ONE. 3 (2): e1565. doi:10,1371 / journal.pone.0001565. PMC 2215775. PMID 18270562.
- ^ Sargent, C. Y .; Berguig, G. Y .; McDevitt, T. C. (2009). „Kardiomyogenní diferenciace embryoidních těl je podporována rotační orbitální suspenzní kulturou“. Tkáňové inženýrství, část A. 15 (2): 331–342. doi:10.1089 / ten. Čaj.2008.0145. PMID 19193130.
- ^ Bauwens, C. L. L .; Peerani, R .; Niebruegge, S .; Woodhouse, K. A .; Kumacheva, E .; Husain, M .; Zandstra, P. W. (2008). „Kontrola kolonie lidských embryonálních kmenových buněk a heterogenita agregátní velikosti ovlivňuje diferenciační trajektorie“. Kmenové buňky. 26 (9): 2300–2310. doi:10.1634 / kmenové články. 2008-0183. PMID 18583540.
- ^ Chen, Y .; Li, X .; Eswarakumar, V. P .; Seger, R .; Lonai, P. (2000). „Fibroblastový růstový faktor (FGF) signalizující prostřednictvím PI 3-kinázy a Akt / PKB je nutný pro diferenciaci embryoidního těla“. Onkogen. 19 (33): 3750–3756. doi:10.1038 / sj.onc.1203726. PMID 10949929.
- ^ Esner, M .; Pachernik, J .; Hampl, A .; Dvořák, P. (2002). „Cílené narušení receptoru 1 fibroblastového růstového faktoru blokuje zrání viscerálního endodermu a kavitaci v myších embryoidních tělech“. International Journal of Developmental Biology. 46 (6): 817–825. PMID 12382948.
- ^ Wan, Y. J .; Wu, T. C .; Chung, A.E .; Damjanov, I. (1984). „Monoklonální protilátky proti lamininu odhalují heterogenitu bazálních membrán ve vývojových a dospělých myších tkáních“. The Journal of Cell Biology. 98 (3): 971–979. doi:10.1083 / jcb.98.3.971. PMC 2113154. PMID 6365932.
- ^ Li, X .; Chen, Y .; Schéele, S .; Arman, E .; Haffner-Krausz, R .; Ekblom, P .; Lonai, P. (2001). „Signalizace růstového faktoru fibroblastů a sestava bazální membrány jsou spojeny během epiteliální morfogeneze těla embrya“. The Journal of Cell Biology. 153 (4): 811–822. doi:10.1083 / jcb.153.4.811. PMC 2192393. PMID 11352941.
- ^ Coucouvanis, E .; Martin, G. R. (1995). „Signály pro smrt a přežití: Dvoustupňový mechanismus kavitace v embryu obratlovců“. Buňka. 83 (2): 279–287. doi:10.1016/0092-8674(95)90169-8. PMID 7585945.
- ^ Smyth, N .; Vatansever, H. S .; Murray, P .; Meyer, M .; Frie, C .; Paulsson, M .; Edgar, D. (1999). „Absence bazálních membrán po cílení na gen LAMC1 vede k embryonální letalitě v důsledku selhání diferenciace endodermu“. The Journal of Cell Biology. 144 (1): 151–160. doi:10.1083 / jcb.144.1.151. PMC 2148127. PMID 9885251.
- ^ Murray, P .; Edgar, D. (2000). „Regulace programované buněčné smrti bazálními membránami v embryonálním vývoji“. The Journal of Cell Biology. 150 (5): 1215–1221. doi:10.1083 / jcb.150.5.1215. PMC 2175256. PMID 10974008.
- ^ Ying, Q. L .; Smith, A. G. (2003). Definované podmínky pro neurální závazek a diferenciaci. Metody v enzymologii. 365. 327–341. doi:10.1016 / s0076-6879 (03) 65023-8. ISBN 9780121822682. PMID 14696356.
- ^ Wiles, M. V .; Keller, G. (1991). "Mnohočetné hematopoetické linie se vyvíjejí z embryonálních kmenových (ES) buněk v kultuře". Rozvoj. 111 (2): 259–267. PMID 1893864.
- ^ Purpura, K. A .; Morin, J .; Zandstra, P. W. (2008). „Analýza časových a na koncentraci závislých účinků BMP-4, VEGF a TPO na vývoj mezodermu a progenitorů krve odvozených z embryonálních kmenových buněk v definovaném médiu bez séra“. Experimentální hematologie. 36 (9): 1186–1198. doi:10.1016 / j.exphem.2008.04.003. PMID 18550259.
- ^ Nostro, M. C .; Cheng, X .; Keller, G. M .; Gadue, P. (2008). „Signalizace Wnt, aktivin a BMP reguluje odlišná stádia vývojové cesty od embryonálních kmenových buněk ke krvi“. Buňková kmenová buňka. 2 (1): 60–71. doi:10.1016 / j.stem.2007.10.011. PMC 2533280. PMID 18371422.
- ^ Ten Berge, D .; Koole, W .; Fuerer, C .; Fish, M .; Eroglu, E .; Nusse, R. (2008). „Wnt Signaling Mediates Self-Organisation and Axis Formation in Embryoid Bodies“. Buňková kmenová buňka. 3 (5): 508–518. doi:10.1016 / j.stem.2008.09.013. PMC 2683270. PMID 18983966.
- ^ Shukla, S .; Nair, R .; Rolle, M. W .; Braun, K. R .; Chan, C. K .; Johnson, P. Y .; Wight, T. N .; McDevitt, T. C. (2009). "Syntéza a organizace hyaluronanu a versikanu embryonálními kmenovými buňkami podstupujícími diferenciaci embryonálních těl". Journal of Histochemistry and Cytochemistry. 58 (4): 345–358. doi:10.1369 / jhc.2009,954826. PMC 2842597. PMID 20026669.
- ^ Bauwens, C.L .; Song, H .; Thavandiran, N .; Ungrin, M .; Massé, S. P .; Nanthakumar, K .; Seguin, C .; Zandstra, P. W. (2011). "Geometrická kontrola kardiomyogenní indukce v lidských pluripotentních kmenových buňkách". Tkáňové inženýrství, část A. 17 (15–16): 1901–1909. doi:10.1089 / ten.TEA.2010.0563. PMID 21417693. S2CID 22010083.
- ^ Eiraku, M .; Takata, N .; Ishibashi, H .; Kawada, M .; Sakakura, E .; Okuda, S .; Sekiguchi, K .; Adachi, T .; Sasai, Y. (2011). „Samoorganizující se morfogeneze optického kalíšku v trojrozměrné kultuře“. Příroda. 472 (7341): 51–56. doi:10.1038 / nature09941. PMID 21475194.
- ^ Bielinska, M .; Narita, N .; Wilson, D. B. (1999). "Zřetelné role pro viscerální endoderm během vývoje embryonálních myší". International Journal of Developmental Biology. 43 (3): 183–205. PMID 10410899.
- ^ Burdsal, C. A .; Damsky, C. H .; Pedersen, R. A. (1993). „Role E-kadherinu a integrinů v mezodermální diferenciaci a migraci na primitivním pruhu savců“. Rozvoj. 118 (3): 829–844. PMID 7521282.
- ^ Finley, K. R .; Tennessen, J .; Shawlot, W. (2003). „Myší vylučovaný frizzlovaný proteinový gen 5 je exprimován v přední viscerální endoderm a endoderm přední části břicha během raného postimplantačního vývoje“. Vzory genového výrazu. 3 (5): 681–684. doi:10.1016 / s1567-133x (03) 00091-7. PMID 12972006.
- ^ Kemp, C .; Willems, E .; Abdo, S .; Lambiv, L .; Leyns, L. (2005). "Exprese všech genů Wnt a jejich vylučovaných antagonistů během vývoje myších blastocyst a postimplantace". Dynamika vývoje. 233 (3): 1064–1075. doi:10.1002 / dvdy.20408. PMID 15880404.
- ^ Rivera-Pérez, J. A .; Magnuson, T. (2005). „Primitivní tvorbě pruhů u myší předchází lokalizovaná aktivace Brachyury a Wnt3.“ Vývojová biologie. 288 (2): 363–371. doi:10.1016 / j.ydbio.2005.09.012. PMID 16289026.
- ^ A b Turner, David; Alonso-Crisostomo, Luz; Girgin, Mehmet; Baillie-Johnson, Peter; Glodowski, Cherise R .; Hayward, Penelope C .; Collignon, Jérôme; Gustavsen, Carsten; Serup, Palle (2017-01-31). „Gastruloidy rozvíjejí tři tělesné osy v nepřítomnosti extraembryonálních tkání a prostorově lokalizované signalizace“. bioRxiv 10.1101/104539.
- ^ A b Turner, David Andrew; Glodowski, Cherise R .; Luz, Alonso-Crisostomo; Baillie-Johnson, Peter; Hayward, Penny C .; Collignon, Jérôme; Gustavsen, Carsten; Serup, Palle; Schröter, Christian (2016-05-13). "Interakce mezi nodální a Wnt signalizací vedou k robustnímu rozbití symetrie a axiální organizaci u gastruloidů (embryonálních organoidů)". bioRxiv 10.1101/051722.
- ^ A b Baillie-Johnson, Peter; Brink, Susanne Carina van den; Balayo, Tina; Turner, David Andrew; Arias, Alfonso Martinez (2015-11-24). „Generování agregátů myších embryonálních kmenových buněk, které vykazují porušení symetrie, polarizaci a vznikající kolektivní chování In Vitro". Žurnál vizualizovaných experimentů (105). doi:10.3791/53252. ISSN 1940-087X. PMC 4692741. PMID 26650833.
- ^ Brink, Susanne C. van den; Baillie-Johnson, Peter; Balayo, Tina; Hadjantonakis, Anna-Katerina; Nowotschin, Sonja; Turner, David A .; Arias, Alfonso Martinez (15. 11. 2014). "Lámání symetrie, specifikace zárodečné vrstvy a axiální organizace v agregátech myších embryonálních kmenových buněk". Rozvoj. 141 (22): 4231–4242. doi:10.1242 / dev.113001. ISSN 0950-1991. PMC 4302915. PMID 25371360.
- ^ Turner, David A .; Hayward, Penelope C .; Baillie-Johnson, Peter; Rué, Pau; Broome, Rebecca; Faunes, Fernando; Arias, Alfonso Martinez (15. 11. 2014). „Signalizace Wnt / β-katenin a FGF řídí specifikaci a údržbu neuromesodermálního axiálního předka v souborech myších embryonálních kmenových buněk“. Rozvoj. 141 (22): 4243–4253. doi:10.1242 / dev.112979. ISSN 0950-1991. PMC 4302903. PMID 25371361.
- ^ A b Kinney, M. A .; Sargent, C. Y .; McDevitt, T. C. (2011). „Multiparametrické účinky hydrodynamických prostředí na kulturu kmenových buněk“. Část B tkáňového inženýrství: Recenze. 17 (4): 249–262. doi:10.1089 / ten.TEB.2011.0040. PMC 3142632. PMID 21491967.
- ^ A b Carpenedo, R. L .; Bratt-Leal, A. S. M .; Marklein, R. A .; Seaman, S. A .; Bowen, N.J .; McDonald, J. F .; McDevitt, T. C. (2009). „Homogenní a organizovaná diferenciace uvnitř embryoidních těl indukovaná mikrosférou zprostředkovanou dodávkou malých molekul“. Biomateriály. 30 (13): 2507–2515. doi:10.1016 / j.biomaterials.2009.01.007. PMC 2921510. PMID 19162317.
- ^ Sachlos, E .; Auguste, D. T. (2008). „Morfologie těla embrya ovlivňuje difúzní transport indukčních biochemikálií: Strategie pro diferenciaci kmenových buněk“. Biomateriály. 29 (34): 4471–4480. doi:10.1016 / j.biomaterials.2008.08.012. PMID 18793799.
- ^ Van Winkle, A. P .; Gates, I.D .; Kallos, M. S. (2012). "Omezení přenosu hmoty v tělech embrya během diferenciace lidských embryonálních kmenových buněk". Buňky Tkáňové orgány. 196 (1): 34–47. doi:10.1159/000330691. PMID 22249133.
- ^ Bratt-Leal, A. S. M .; Carpenedo, R. L .; Ungrin, M. D .; Zandstra, P. W .; McDevitt, T. C. (2011). „Začlenění biomateriálů do mnohobuněčných agregátů moduluje diferenciaci pluripotentní kmenové buňky“. Biomateriály. 32 (1): 48–56. doi:10.1016 / j.biomaterials.2010.08.113. PMC 2987521. PMID 20864164.
- ^ Purpura, K. A .; Bratt-Leal, A. S. M .; Hammersmith, K. A .; McDevitt, T. C .; Zandstra, P. W. (2012). „Systematické inženýrství 3D pluripotentních výklenků kmenových buněk pro vedení vývoje krve“. Biomateriály. 33 (5): 1271–1280. doi:10.1016 / j.biomaterials.2011.10.051. PMC 4280365. PMID 22079776.
- ^ Bratt-Leal, A. S. M .; Kepple, K. L .; Carpenedo, R. L .; Cooke, M. T .; McDevitt, T. C. (2011). „Magnetická manipulace a prostorové vzorkování agregátů mnohobuněčných kmenových buněk“. Integrativní biologie. 3 (12): 1224–1232. doi:10.1039 / c1ib00064k. PMC 4633527. PMID 22076329.
- ^ Akins, R.E .; Rockwood, D .; Robinson, K. G .; Sandusky, D .; Rabolt, J .; Pizarro, C. (2010). „Trojrozměrná kultura mění primární fenotyp srdečních buněk“. Tkáňové inženýrství, část A. 16 (2): 629–641. doi:10.1089 / ten.tea.2009.0458. PMC 2813151. PMID 20001738.
- ^ Chang, T. T .; Hughes-Fulford, M. (2009). „Monovrstva a sféroidní kultura buněk jaterních buněk hepatocelulárního karcinomu jater prokazuje odlišné vzory globálních expresí a funkční fenotypy“. Tkáňové inženýrství, část A. 15 (3): 559–567. doi:10.1089 / ten. Čaj.2007.0434. PMC 6468949. PMID 18724832.