Autapomorphy - Autapomorphy
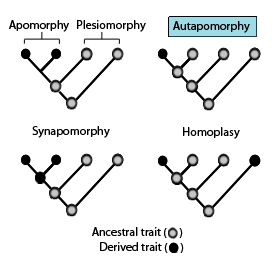
v fylogenetika, an autapomorphy je charakteristický rys, známý jako odvozený vlastnost, která je pro daný taxon jedinečná. To znamená, že se nachází pouze v jednom taxon, ale nebyl nalezen v žádném jiném nebo outgroup taxony, dokonce ani ty, které nejblíže souvisejí s fokálním taxonem (což může být druh, rodina nebo obecně jakýkoli clade).[2] Lze jej proto považovat za apomorfie ve vztahu k jedinému taxonu.[3] Slovo autapomorphy, poprvé představen v roce 1950 německy entomolog Willi Hennig, je odvozen z řecký slova αὐτός, aut- = "já"; ,πό, apo = "od"; a μορφή, morphḗ = "tvar".[4]
Diskuse
Protože autapomorfie jsou přítomny pouze v jediném taxonu, nepřenášejí informace o vztahu. Autapomorfie proto nejsou užitečné k odvození fylogenetických vztahů. Autapomorphy, jako synapomorfie a plesiomorphy je relativní pojem v závislosti na daném taxonu. Autapomorfie na dané úrovni může být synapomorfie na méně inkluzivní úrovni.[5] Příklad autapomorfie lze popsat u moderních hadů. Hadi ztratili dva páry nohou, které charakterizují všechny Tetrapoda a nejbližší taxony k Ophidia - stejně jako jejich společní předkové - všichni mají dva páry nohou. Proto taxon Ophidia představuje autapomorfii s ohledem na absenci nohou.[3]
The koncept autapomorfních druhů je jednou z mnoha metod, které vědci mohou použít k definování a rozlišení druh od sebe navzájem. Tato definice přiřazuje druhy na základě míry divergence spojené s reprodukční nekompatibilitou, která se měří v podstatě počtem autapomorfií.[6] Tato metoda seskupování se často označuje jako „monofyletický koncept druhu "nebo koncept" phylospecies "a popularizoval jej D.E. Rosen v roce 1979. V rámci této definice je druh považován za" nejméně inkluzivní monofyletickou skupinu definovatelnou alespoň jednou autapomorfií ".[7] I když je tento model speciace užitečný v tom, že se vyhýbá nemonofyletickým seskupením, má také svou kritiku. N.I. Platnick například věří, že koncept autapomorfního druhu je neadekvátní, protože umožňuje možnost reprodukční izolace a speciace a zároveň zruší „druhový“ stav mateřské populace. Jinými slovy, pokud se periferní populace odtrhne a stane se reprodukčně izolovanou, bude pravděpodobně nutné vyvinout alespoň jednu autapomorfii, aby byla uznána jako jiný druh. Pokud se to může stát bez větší mateřské populace taky při vývoji nové autapomorfie pak mateřská populace nemůže zůstat druhem v rámci konceptu autapomorfního druhu: již by neměla žádné apomorfie, které by nesdílely ani dceřinné druhy.[8]
Fylogenetické podobnosti: Tyto fylogenetické termíny se používají k popisu různých vzorů stavů předků a odvozených znaků nebo vlastností, jak je uvedeno ve výše uvedeném diagramu ve spojení se synapomorfiemi.[1]
- Homoplasy v biologická systematika je, když a vlastnost byl během evoluce získán nebo ztracen nezávisle v samostatných liniích. Tento konvergentní evoluce vede k tomu, že druhy nezávisle sdílejí rys, který se liší od rysu, o kterém se usuzuje, že byl přítomen u jejich společného předka.[9][10][11]
- Paralelní Homoplasy - odvozený znak přítomný ve dvou skupinách nebo druzích bez společného předka kvůli konvergentní evoluce.[12]
- Zvrátit Homoplasy - vlastnost přítomná u předka, ale ne u přímých potomků, která se znovu objeví u pozdějších potomků.[13]
- Apomorfie - odvozená vlastnost. Apomorphy sdílená dvěma nebo více taxony a zděděná od společného předka je synapomorphy. Apomorfie jedinečná pro daný taxon je autapomorfie.[14][15][16][17]
- Synapomorphy /Homologie - odvozený znak, který se nachází v některých nebo všech koncových skupinách kladu a zdědil se od společného předka, pro který byl autapomorfií (tj. své bezprostřední předek).
- Základní synapomorfie - synapomorfie, která se opět ztratila u mnoha členů kladu. Pokud se ztratí u všech kromě jedné, může být těžké ji odlišit od autapomorfie.
- Autapomorphy - výrazný odvozený znak, který je jedinečný pro daný taxon nebo skupinu.[3]
- Symptomiomorphy - vlastnost předků sdílená dvěma nebo více taxony.
- Plesiomorphy - symplesiomorfie diskutovaná v odkazu na odvozenější stav.
- Pseudoplesiomorphy - je vlastnost, kterou nelze identifikovat ani jako plesiomorphy, ani jako apomorfie, která je obrácením.[18]
- Zvrat - je ztráta odvozeného znaku přítomného u předka a obnovení plesiomorfního znaku.
- Konvergence - nezávislý vývoj podobného znaku u dvou nebo více taxonů.
- Hemiplasy[19][20]
Reference
- ^ A b Page RD, Holmes EC (14. července 2009). Molekulární evoluce: fylogenetický přístup. John Wiley & Sons. ISBN 978-1-4443-1336-9. OCLC 609843839.
- ^ Futuyma DJ (1998). Evoluční biologie (3. vyd.). Sinauer Associates, Inc. str. 95.
- ^ A b C Appel RD, Feytmans E (2009). „Kapitola 3: Úvod do fylogenetiky a její molekulární aspekty“. Bioinformatika: švýcarská perspektiva (1. vyd.). Světová vědecká nakladatelská společnost.
- ^ Calow PP (2009). Encyklopedie ekologie a environmentálního managementu. John Wiley & Sons. ISBN 978-1-4443-1324-6. OCLC 1039167559.
- ^ Forey PL (1997). Historie Coelacanth ryb (1. vyd.). Sprinter.
- ^ Howard DJ, Berlocher SH (1998). Nekonečné formy: druhy a speciace (1. vyd.). USA: Oxford University Press. ISBN 978-0-19-510901-6. OCLC 60181901.
- ^ Bull AT (2004). Mikrobiální rozmanitost a biologický průzkum. Stiskněte ASM.
- ^ Platnick NI (2001). "Od kladogramů po klasifikace: Cesta k dePhylocode." (PDF). Systematické sdružení.
- ^ Gauger A (17. dubna 2012). „Podobnost se stává! Problém homoplasie“. Evolution Today & Science News.
- ^ Sanderson MJ, Hufford L (21. října 1996). Homoplasy: Opakování podobnosti v evoluci. Elsevier. ISBN 978-0-08-053411-4. OCLC 173520205.
- ^ Brandley MC, Warren DL, Leaché AD, McGuire JA (duben 2009). "Homoplasy and clade support". Systematická biologie. 58 (2): 184–98. doi:10.1093 / sysbio / syp019. PMID 20525577.
- ^ Archie JW (září 1989). „Homoplasy Excess Ratios: New Indices for Measuring Levels of Homoplasy in Phylogenetic Systematics and a Critique of the Consistency Index“. Systematická zoologie. 38 (3): 253–269. doi:10.2307/2992286. JSTOR 2992286.
- ^ Wake DB, Wake MH, Specht CD (únor 2011). "Homoplasy: od detekčního vzorce k určování procesu a mechanismu evoluce". Věda. 331 (6020): 1032–5. doi:10.1126 / science.1188545. PMID 21350170. S2CID 26845473. Shrnutí ležel – Věda denně.
- ^ Simpson MG (9. srpna 2011). Systematika rostlin. Elsevier. Amsterdam: Elsevier. ISBN 9780080514048.
- ^ Russell PJ, Hertz PE, McMillan B (2013). Biology: The Dynamic Science. Cengage Learning. ISBN 978-1-285-41534-5.
- ^ Lipscomb D (1998). "Základy kladistické analýzy" (PDF). Washington D.C .: Univerzita George Washingtona.
- ^ Choudhuri S (09.05.2014). Bioinformatika pro začátečníky: geny, genomy, molekulární evoluce, databáze a analytické nástroje (1. vyd.). Akademický tisk. str. 51. ISBN 978-0-12-410471-6. OCLC 950546876.
- ^ Williams D, Schmitt M, Wheeler Q (2016-07-21). Budoucnost fylogenetické systematiky: Dědictví Williho Henniga. ISBN 978-1-107-11764-8. OCLC 951563305.
- ^ Avise JC, Robinson TJ (červen 2008). „Hemiplasy: nový termín ve slovníku fylogenetiky“. Systematická biologie. 57 (3): 503–7. doi:10.1080/10635150802164587. PMID 18570042.
- ^ Copetti D, Búrquez A, Bustamante E, Charboneau JL, Childs KL, Eguiarte LE a kol. (Listopad 2017). „Rozsáhlá diskordance genového stromu a hemiplasie formovaly genomy severoamerických sloupovitých kaktusů“. Sborník Národní akademie věd Spojených států amerických. 114 (45): 12003–12008. doi:10.1073 / pnas.1706367114. PMC 5692538. PMID 29078296.
| Relevantní pole | ||
|---|---|---|
| Základní pojmy | ||
| Inferenční metody | ||
| Aktuální témata | ||
| Skupinové vlastnosti | ||
| Skupinové typy | ||
| Nomenklatura | ||
| ||