Cyklol - Cyclol - Wikipedia
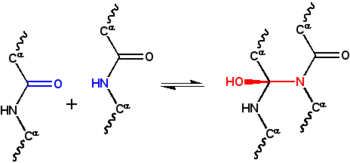
The cyklol hypotéza je první strukturální model a složený, kulovitý protein.[1] Byl vyvinut společností Dorothy Wrinch na konci třicátých let a byla založena na třech předpokladech. Za prvé, hypotéza předpokládá, že dva peptidové skupiny lze zesíťovat pomocí a cyklol reakce (Obrázek 1); tyto křížové odkazy jsou kovalentní analogy nekovalentní Vodíkové vazby mezi peptidovými skupinami. Tyto reakce byly pozorovány v ergopeptidy a další sloučeniny. Za druhé předpokládá, že za určitých podmínek aminokyseliny přirozeně vytvoří maximální možný počet cyklických zesíťování, což povede k molekuly cyklolu (Obrázek 2) a cyklol tkaniny (Obrázek 3). Tyto molekuly a látky cyklolu nebyly nikdy pozorovány. Nakonec hypotéza předpokládá, že globulární proteiny mají a terciární struktura souhlasí s Platonické pevné látky a semiregular polyhedra vytvořeno z cyklolových tkanin bez volných hran. Také takové molekuly "uzavřeného cyklolu" nebyly pozorovány.
Ačkoli pozdější údaje prokázaly, že tento původní model pro struktura globulárních proteinů bylo třeba upravit, bylo ověřeno několik prvků cyklolového modelu, například samotná cyklolová reakce a hypotéza, že hydrofobní interakce jsou zodpovědní hlavně za skládání bílkovin. Cyklická hypotéza stimulovala mnoho vědců k výzkumu otázek ve struktuře proteinů a chemii a byla předchůdcem přesnějších modelů předpokládaných pro DNA dvojitá šroubovice a sekundární struktura bílkovin. Návrh a testování modelu cyklol také poskytuje vynikající ilustraci empirický padělání jednající jako součást vědecká metoda.
Historický kontext
Do poloviny 30. let analytická ultracentrifugace studie od Theodor Svedberg ukázaly, že proteiny mají dobře definovanou chemickou strukturu a nejedná se o agregace malých molekul.[2] Stejné studie ukázaly, že molekulová hmotnost proteinů spadala do několika dobře definovaných tříd souvisejících s celými čísly,[3] jako Mw = 2p3q Da, kde p a q jsou nezáporná celá čísla.[4] Bylo však obtížné určit přesnou molekulovou hmotnost a počet aminokyselin v proteinu. Svedberg také ukázal, že změna podmínek v roztoku může způsobit rozložení proteinu na malé podjednotky, nyní známé jako změna kvartérní struktura.[5]
The chemická struktura z bílkoviny byl v té době ještě debatován.[6] Nejpřijatelnější (a nakonec správnou) hypotézou bylo, že proteiny jsou lineární polypeptidy, tj. nerozvětvený polymery z aminokyseliny propojeno uživatelem peptidové vazby.[7][8] Typický protein je však pozoruhodně dlouhý - stovky aminokyselinové zbytky —A několik významných vědců si nebylo jistých, zda jsou tak dlouhé, lineární makromolekuly může být stabilní v řešení.[9][10] Další pochybnosti o polypeptidová povaha proteinů vznikl proto, že někteří enzymy Bylo pozorováno, že štěpí proteiny, ale ne peptidy, zatímco jiné enzymy štěpí peptidy, ale ne složené proteiny.[11] Pokusy o syntézu proteinů ve zkumavce byly neúspěšné, hlavně kvůli chirality aminokyselin; přirozeně se vyskytující proteiny jsou složeny pouze z levák aminokyseliny. Zvažovaly se proto alternativní chemické modely proteinů, jako je hypotéza diketopiperazinu z Emil Abderhalden.[12][13] Žádný alternativní model však dosud nevysvětlil, proč proteiny po hydrolýze a proteolýze poskytují pouze aminokyseliny a peptidy. Jak objasnil Linderstrøm-Lang,[14] tyto proteolytické údaje to ukázaly denaturované proteiny byly polypeptidy, ale dosud nebyla získána žádná data o struktuře skládaných proteinů; denaturace tedy může zahrnovat chemickou změnu, která přeměňuje skládané proteiny na polypeptidy.
Proces bílkovin denaturace (na rozdíl od koagulace ) objevil v roce 1910 Harriette Chick a Charles Martin,[15] ale jeho povaha byla stále záhadná. Tim Anson a Alfred Mirsky ukázal, že denaturace byla reverzibilní, dvoustavový proces[16] což má za následek dostupnost mnoha chemických skupin pro chemické reakce, včetně štěpení enzymy.[17] V roce 1929 Hsien Wu předpokládal správně, že denaturace odpovídala odvíjení proteinu, čistě konformační změně, která vedla k vystavení aminokyselinových postranních řetězců rozpouštědlu.[18] Wuova hypotéza byla rovněž nezávisle předložena v roce 1936 Mirským a Linus Pauling.[19] Vědci o bílkovinách nicméně nemohli vyloučit možnost, že denaturace odpovídá a chemikálie změna ve struktuře bílkovin,[17] hypotéza, která byla až do padesátých let považována za (vzdálenou) možnost.[20][21]
Rentgenová krystalografie právě začala jako disciplína v roce 1911 a poměrně rychle postupovala od jednoduchých krystalů solí k krystalům komplexních molekul, jako jsou cholesterol. I ty nejmenší proteiny však mají přes 1 000 atomů, což činí jejich strukturu mnohem složitější. V roce 1934 Dorothy Crowfoot Hodgkin vzal krystalografická data o struktuře malého proteinu, inzulín, i když struktura tohoto a dalších proteinů byla vyřešena až koncem šedesátých let. Průkopnické rentgen difrakce vláken údaje byly shromážděny na počátku 30. let pro mnoho přirozených vláknité proteiny jako je vlna a vlasy William Astbury, který navrhl primitivní modely sekundární struktura prvky jako alfa šroubovice a beta list.
Od té doby proteinová struktura byl ve 30. letech tak špatně pochopen, nebyly známy ani fyzické interakce odpovědné za stabilizaci této struktury. Astbury předpokládal, že struktura vláknité proteiny byl stabilizován Vodíkové vazby v β-listech.[22][23] Myšlenka, že globulární proteiny jsou také stabilizovány vodíkovými vazbami navrhl Dorothy Jordan Lloyd[24][25] v roce 1932 a později prosazován Alfred Mirsky a Linus Pauling.[19] Na přednášce Astburyho z roku 1933 před Oxfordskou vědeckou společností, fyzikem Frederick Frank navrhl, že vláknitý protein α-keratin by mohl být stabilizován alternativním mechanismem, jmenovitě, kovalentní síťování peptidové vazby cyklickou reakcí výše.[26] Cyklické síťování přitahuje dvě peptidové skupiny blízko sebe; atomy N a C jsou odděleny ~ 1,5A, zatímco jsou odděleny ~ 3A v typickém vodíková vazba. Ta myšlenka zaujala J. D. Bernal, který to navrhl matematikovi Dorothy Wrinch jako možná užitečné pro pochopení struktury proteinů.
Základní teorie
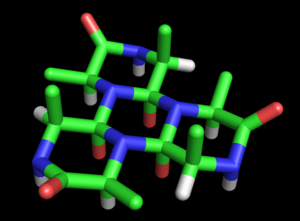
Wrinch rozvinul tento návrh do plnohodnotného modelu proteinová struktura. Základní model cyklolu byl uveden v její první práci (1936).[27] Všimla si možnosti, že polypeptidy může cyklizovat za vzniku uzavřených kruhů (skutečný ) a že tyto kruhy mohou tvořit vnitřní síťování prostřednictvím cyklické reakce (také pravdivé, i když vzácné). Za předpokladu, že cyklol forma peptidová vazba může být stabilnější než amidová forma, Wrinch dospěl k závěru, že určité cyklické peptidy by přirozeně vytvořil maximální počet cyklolových vazeb (např cyklol 6, Obrázek 2). Takové molekuly cyklolu by měly hexagonální symetrii, kdyby chemické vazby byly brány jako mající stejnou délku, zhruba 1,5A; pro srovnání mají vazby N-C a C-C délky 1,42 Á, respektive 1,54 Á.
Tyto kroužky lze prodloužit na neurčito, aby vytvořily a cyklol tkanina (Obrázek 3). Takové látky vykazují kvazi-krystalický řád s dlouhým dosahem, který Wrinch cítil pravděpodobně v proteinech, protože musí hustě zabalit stovky zbytků. Další zajímavou vlastností takových molekul a tkanin je, že jejich aminokyselina postranní řetězy směřují axiálně nahoru pouze z jedné strany; opačný obličej nemá postranní řetězy. Jedna tvář je tedy zcela nezávislá na primární sekvence peptidu, který Wrinch předpokládal, by mohl odpovídat za vlastnosti proteinů nezávislé na sekvenci.
Ve svém původním článku Wrinch jasně uvedla, že model cyklol byl pouze a pracovní hypotéza, potenciálně platný model proteinů, který by musel být zkontrolován. Jejími cíli v tomto článku a jeho nástupcích bylo navrhnout dobře definovaný testovatelný model, vypracovat důsledky jeho předpokladů a předpovědi, které by mohly být experimentálně testovány. V těchto cílech uspěla; experimenty a další modelování však během několika let ukázaly, že hypotéza o cyklolu je jako model globulárních proteinů neudržitelná.
Stabilizující energie

Ve dvou tandemových Dopisech redaktorovi (1936),[28][29] Wrinch a Frank se zabývali otázkou, zda je cyklolová forma peptidové skupiny skutečně stabilnější než forma amidová. Relativně jednoduchý výpočet ukázal, že forma cyklolu je významně méně stabilní než amidová forma. Cyklický model by proto musel být opuštěn, pokud by nebyl identifikován kompenzační zdroj energie. Frank původně navrhoval, aby se forma cyklolu mohla stabilizovat lepšími interakcemi s okolním rozpouštědlem; později Wrinch a Irving Langmuir Předpokládá se, že hydrofobní asociace nepolárních postranních řetězců poskytuje stabilizační energii k překonání energetické ceny cyklických reakcí.[30][31]
Labilita cyklolové vazby byla považována za výhoda modelu, protože poskytoval přirozené vysvětlení vlastností denaturace; reverze cyklolových vazeb na jejich stabilnější amidovou formu by otevřela strukturu a umožnila útokům na tyto vazby proteázy, v souladu s experimentem.[32][33] Rané studie ukázaly, že bílkoviny denaturovaný podle tlak jsou často v jiném stavu než stejné proteiny denaturované vysokou teplota, který byl interpretován jako možná podporující cyklol model denaturace.[34]
Langmuir-Wrinchova hypotéza hydrofobní stabilizace se podílela na pádu cyklolového modelu, hlavně kvůli vlivu Linus Pauling, kteří podporovali hypotézu, že proteinová struktura byla stabilizována Vodíkové vazby. Muselo uplynout dalších dvacet let, než byly hydrofobní interakce uznány jako hlavní hnací síla skládání proteinů.[35]
Sterická komplementarita
Ve svém třetím příspěvku o cyklolech (1936)[36] Wrinch poznamenal, že mnoho "fyziologicky aktivních" látek, jako je steroidy jsou složeny z kondenzovaných šestihranných kruhů atomů uhlíku, a proto by mohly být stericky komplementární do tváře molekul cyklolu bez aminokyselina boční řetězy. Wrinch navrhl, že sterická komplementarita je jedním z hlavních faktorů při určování, zda se malá molekula váže na protein.
Wrinch spekuloval, že proteiny jsou zodpovědné za syntézu všech biologických molekul. S vědomím, že buňky tráví své bílkoviny pouze za extrémních hladových podmínek, Wrinch dále spekuloval, že život by bez bílkovin nemohl existovat.
Hybridní modely
Od začátku byla cyklolová reakce považována za kovalentní analog sloučeniny vodíková vazba. Bylo proto přirozené uvažovat o hybridních modelech s oběma typy dluhopisů. To bylo předmětem čtvrtého Wrinchova článku o modelu cyklol (1936),[37] napsáno společně s Dorothy Jordan Lloyd, který nejprve navrhl, aby byly globulární proteiny stabilizovány vodíkovými vazbami.[24] V roce 1937 byl napsán navazující článek, který odkazoval na další výzkumníky týkající se vodíkových vazeb v proteinech, jako např Maurice Loyal Huggins a Linus Pauling.[38]
Wrinch také napsal příspěvek s William Astbury s poukazem na možnost keto-enol izomerizace> CαHα a amidovou karbonylovou skupinu> C = O, produkující zesíťování> Cα-C (OHα) [39] Takové reakce by mohly poskytnout pětičlenné kruhy, zatímco klasická cyklolová hypotéza vytváří šestičlenné kruhy. Tato hypotéza keto-enolového zesítění nebyla vyvinuta mnohem dále.[40]
Prostorové textilie

Ve své páté práci o cyklolech (1937),[41] Wrinch identifikoval podmínky, za kterých lze spojit dvě rovinné cyklolové látky, aby vytvořily úhel mezi svými rovinami při respektování úhlů chemických vazeb. Identifikovala matematické zjednodušení, ve kterém mohou být nerovinné šestičlenné kruhy atomů reprezentovány rovinnými "středními šestiúhelníky" vytvořenými ze středů chemických vazeb. Toto znázornění "středního šestiúhelníku" usnadnilo pochopení toho, že roviny cyklolové tkaniny lze spojit správně, pokud vzepětí úhel mezi rovinami se rovná čtyřbokému vazebnému úhlu δ = arccos (-1/3) ≈ 109,47 °.
Lze sestavit širokou škálu uzavřených mnohostěnů splňujících toto kritérium, z nichž nejjednodušší jsou zkrácený čtyřstěn, zkrácený osmistěn a osmistěn, což jsou Platonické pevné látky nebo semiregular polyhedra. Když vezmeme v úvahu první sérii „uzavřených cyklolů“ (modelových na zkráceném čtyřstěnu), Wrinch ukázal, že jejich počet aminokyselin zvýšil kvadraticky jako 72n2, kde n je index uzavřeného cyklolu Cn. To znamená, že C1 cyklol má 72 zbytků, C2 cyklol má 288 zbytků atd. Předběžná experimentální podpora této predikce pochází z Max Bergmann a Carl Niemann,[4] jehož analýzy aminokyselin naznačují, že proteiny byly složeny z celočíselných násobků 288 aminokyselina zbytky (n= 2). Obecněji řečeno, cyklol model globulárních proteinů představoval počátek analytická ultracentrifugace výsledky Theodor Svedberg, což naznačuje, že molekulové hmotnosti bílkovin spadalo do několika tříd souvisejících s celými čísly.[2][3]
Cyklický model byl v souladu s obecnými vlastnostmi, které se pak přisuzovaly složeným proteinům.[42] (1) Centrifugační studie ukázaly, že složené proteiny byly výrazně hustší než voda (~ 1,4G /ml ), a tedy těsně zabalené; Wrinch předpokládal, že by to mělo znamenat husté balení pravidelný balení. (2) Navzdory své velké velikosti některé proteiny snadno krystalizují do symetrických krystalů, což odpovídá myšlence symetrických ploch, které se při asociaci shodují. (3) Bílkoviny váží ionty kovů; protože kovová vazebná místa musí mít specifickou geometrii vazby (např. oktaedrickou), bylo možné předpokládat, že celý protein měl také podobně krystalickou geometrii. (4) Jak je popsáno výše, model cyklol poskytl jednoduchý chemikálie vysvětlení denaturace a obtížnost štěpení složených proteinů proteázami. (5) Předpokládalo se, že proteiny jsou odpovědné za syntézu všech biologických molekul, včetně dalších proteinů. Wrinch poznamenal, že pevná, jednotná struktura by byla užitečná pro proteiny při vytváření vlastní syntézy, analogické k Watson -Francis Crick koncept DNA šablonování vlastní replikace. Vzhledem k tomu, že mnoho biologických molekul, jako je cukry a steroly mají hexagonální strukturu, bylo pravděpodobné předpokládat, že jejich syntetizující proteiny měly také hexagonální strukturu. Wrinch shrnula svůj model a podpůrná experimentální data o molekulové hmotnosti ve třech přehledových článcích.[43]
Předpokládané proteinové struktury
Poté, co Wrinch navrhl model globulárních proteinů, zkoumal, zda je v souladu s dostupnými strukturálními údaji. Předpokládala, že bovinní tuberkulinový protein (523) je a C1 uzavřený cyklol sestávající ze 72 zbytků[44] a to zažívací enzym pepsin byl C2 uzavřený cyklol 288 zbytků.[45][46] Tyto předpovědi počtu reziduí bylo obtížné ověřit, protože metody dostupné k měření hmotnosti proteinů byly nepřesné, jako například analytická ultracentrifugace a chemické metody.
Wrinch to také předpověděl inzulín byl C2 uzavřený cyklol skládající se z 288 zbytků. Pro inzulin byla k dispozici omezená rentgenová krystalografická data, která Wrinch interpretovala jako „potvrzující“ její model.[47] Tato interpretace však vyvolala poměrně tvrdou kritiku za to, že byla předčasná.[48] Pečlivé studie Pattersonových diagramů inzulínu Dorothy Crowfoot Hodgkin ukázal, že jsou zhruba v souladu s cyklolovým modelem; dohoda však nebyla dost dobrá na to, aby tvrdila, že model cyklol byl potvrzen.[49]
Nepravděpodobnost modelu
Ukázalo se, že cyklolová tkanina je nepravděpodobná z několika důvodů. Hans Neurath a Henry Bull prokázali, že husté balení postranních řetězců v cyklolové tkanině bylo nekonzistentní s experimentální hustotou pozorovanou v proteinových filmech.[50] Maurice Huggins vypočítal, že několik nevázaných atomů cyklolové struktury se přiblíží blíže, než dovoluje jejich van der Waalsovy poloměry; například vnitřní Hα a C.α atomy mezer by byly odděleny pouze 1,68A (Obrázek 5).[51] Haurowitz chemicky ukázal, že vnějšek proteinů nemohl mít velké množství hydroxylových skupin, což je klíčová predikce cyklolového modelu,[52] vzhledem k tomu, že Meyer a Hohenemser ukázali, že cyklické kondenzace aminokyselin neexistovaly ani v nepatrném množství jako přechodný stav.[53] Obecnější chemické argumenty proti modelu cyklol uvedli Bergmann a Niemann[54] a tím Neuberger.[55][56] Infračervená spektroskopická data ukázala, že počet karbonylových skupin v proteinu se při hydrolýze nezměnil,[57] a že intaktní skládané proteiny mají úplnou komplement amid karbonylových skupin;[58] obě pozorování jsou v rozporu s cyklolovou hypotézou, že takové karbonyly se převádějí na hydroxylové skupiny ve složených proteinech. Nakonec bylo známo, že proteiny obsahují prolin ve významných množstvích (obvykle 5%); protože prolin postrádá amidový vodík a jeho dusík již tvoří tři kovalentní vazby, zdá se, že prolin není schopen cyklické reakce a nemůže být začleněn do cyklolové struktury. Encyklopedické shrnutí chemických a strukturních důkazů proti cyklol modelu poskytli Pauling a Niemann.[59] Navíc podpůrný důkaz - výsledek, že všechny proteiny obsahují celočíselný násobek 288 aminokyselina zbytky[4]—V roce 1939 se rovněž ukázalo jako nesprávné.[60]
Wrinch odpověděl na kritiku stérického střetu, volné energie, chemikálií a zbytkových čísel cyklolového modelu. U stérických střetů poznamenala, že malé deformace úhlů a délek vazeb by umožnily těmto stérickým střetům ulevit nebo je alespoň snížit na rozumnou úroveň.[61] Poznamenala, že vzdálenosti mezi nevázanými skupinami v rámci jedné molekuly mohou být kratší, než se od nich očekávalo van der Waalsovy poloměry, např. 2.93A vzdálenost mezi methylovými skupinami v hexamethylbenzenu. Pokud jde o pokutu za volnou energii pro cyklolovou reakci, Wrinch nesouhlasil s Paulingovými výpočty a uvedl, že o intramolekulárních energiích je známo příliš málo na to, aby samotný cyklol model byl vyloučen pouze na tomto základě.[61] V reakci na chemickou kritiku Wrinch navrhl, že studované modelové sloučeniny a jednoduché bimolekulární reakce se nemusí týkat cyklolového modelu a že sterická zábrana mohla zabránit reakci povrchových hydroxylových skupin.[62] Na základě kritiky počtu reziduí Wrinch rozšířila svůj model tak, aby umožňoval další počty reziduí. Zejména vytvořila „minimální“ uzavřený cyklol s pouhými 48 zbytky,[63] a na tomto (nesprávném) základě mohl jako první navrhnout, že inzulín monomer měl molekulovou hmotnost zhruba 6000Da.[64][65]
Proto tvrdila, že cyklol model globulárních proteinů je stále potenciálně životaschopný[66][67] a dokonce navrhl cyklolovou tkaninu jako součást cytoskelet.[68] Většina vědců o bílkovinách tomu však přestala věřit a Wrinch obrátila svou vědeckou pozornost na matematické problémy Rentgenová krystalografie, k čemuž významně přispěla. Jedinou výjimkou byl fyzik Gladys Anslow, Wrinchův kolega v Smith College, který studoval ultrafialový vstřebávání spektra proteinů a peptidů ve 40. letech 20. století a umožnila možnost cyklů při interpretaci jejích výsledků.[69][70] Jako sekvence inzulínu začaly být určovány Frederick Sanger, Anslow zveřejnil trojrozměrný cyklol model s bočními řetězci,[71] založený na páteři Wrinchova modelu „minimálního cyklolu“ z roku 1948.[63]
Částečné vykoupení

Pád celkového modelu cyklolu obecně vedl k odmítnutí jeho prvků; jedna významná výjimka byla J. D. Bernal krátkodobé přijetí Langmuir-Wrinchovy hypotézy, že skládání bílkovin je poháněn hydrofobní asociací.[73] Přesto byly cyklolové vazby identifikovány v malých, přirozeně se vyskytujících cyklické peptidy v padesátých letech.
Objasnění moderní terminologie je na místě. Klasická cyklolová reakce je přidání NH aminu a peptidová skupina na C = O karbonylovou skupinu jiného; výsledná sloučenina se nyní nazývá azacyklol. Analogicky, an oxacykl vzniká, když se hydroxylová skupina OH přidá k peptidylové karbonylové skupině. Podobně, a thiacyclol se tvoří přidáním SH thiolové skupiny k peptidylové karbonylové skupině.[74]
Oxacyclol alkaloid ergotamin z houba Claviceps purpurea byl první identifikovaný cyklol.[75] Cyklický depsipeptid serratamolid je také tvořen oxacyklolovou reakcí.[76] Byly také získány chemicky analogické cyklické thiacykloly.[77] Klasické azacykloly byly pozorovány u malých molekul[78] a tripeptidy.[79] Peptidy jsou přirozeně produkovány reverzí azacylolů,[80] klíčová předpověď modelu cyklol. Navzdory tomu byly nyní identifikovány stovky molekul cyklolu Linus Pauling Výpočet, že takové molekuly by neměly existovat kvůli jejich nepříznivě vysoké hodnotě energie.[59]
Po dlouhé přestávce, během které pracovala hlavně na matematice Rentgenová krystalografie Wrinch reagoval na tyto objevy s novým nadšením pro cyklol model a jeho význam v biochemii.[81] Vydala také dvě knihy popisující teorii cyklolu a malé peptidy obecně.[82][83]
Ilustrace vědecké metody
Cyklický model proteinové struktury je příkladem empirický padělání jednající jako součást vědecká metoda. Originál hypotéza je vysvětleno nevysvětlitelné experimentální pozorování; jsou zpracovány důsledky této hypotézy, což vede k předpovědím, které jsou testovány experimentem. V tomto případě byla klíčová hypotéza, že cyklol forma peptidové skupiny může být upřednostňována před amidovou formou. Tato hypotéza vedla k předpovědím molekuly cyklol-6 a cyklolové struktury, což zase navrhlo model polopravidelného mnohostěnu pro globulární proteiny. Klíčovou testovatelnou predikcí bylo, že karbonylové skupiny složeného proteinu by měly být z velké části převedeny na hydroxylové skupiny; spektroskopické a chemické experimenty však ukázaly, že tato předpověď byla nesprávná. Cyklický model také předpovídá vysokou laterální hustotu aminokyselin ve složených proteinech a ve filmech, které nesouhlasí s experimentem. Proto mohl být model cyklol odmítnut a bylo zahájeno hledání nových hypotéz o proteinová struktura, jako jsou modely alfa šroubovice navrženo ve 40. a 50. letech.
Někdy se tvrdí, že cyklická hypotéza nikdy neměla být pokročilá,[84][85] kvůli jeho a priori nedostatky, např. stérické střety, neschopnost přizpůsobit se prolin a vysoká energie zdarma znevýhodnění samotné cyklolové reakce. I když takové nedostatky poskytly cyklolovou hypotézu nepravděpodobný, nezvládli to nemožné. Cyklický model byl první dobře definovanou strukturou navrženou pro globulární proteiny a o intramolekulárních silách a struktuře proteinů bylo tehdy známo příliš málo na to, aby je bylo možné okamžitě odmítnout. Úhledně vysvětlil několik obecných vlastností proteinů a vysvětlil tehdejší anomální experimentální pozorování. Ačkoli obecně nesprávné, některé prvky teorie cyklolu byly nakonec ověřeny, jako jsou reakce cyklolu a role hydrofobní interakce v skládání bílkovin. Užitečné srovnání je Bohrův model z vodík atom, který byl od svého vzniku považován za nepravděpodobný, dokonce i jeho tvůrcem,[86] přesto vedl cestu k nakonec správné teorii kvantová mechanika. Podobně, Linus Pauling navrhla dobře definovaný model DNA[87] to bylo rovněž nepravděpodobné, ale pro ostatní vyšetřovatele provokativní.[88][89]
Naopak model cyklol je příkladem nesprávné vědecké velké teorie symetrie a krása, dvě vlastnosti, které lze považovat za známky „zjevně pravdivých“ vědeckých teorií. Například Watson -Cricku dvojitá spirála model DNA[89] se někdy říká, že je „zřejmý“, protože je pravděpodobný vodíkové vazby a symetrie; za jiných podmínek jsou však upřednostňovány jiné, méně symetrické struktury DNA.[90] Podobně krásná teorie obecná relativita byl považován za Albert Einstein protože nepotřebuje experimentální ověření; ale i tato teorie bude vyžadovat revizi, aby byla konzistentní s kvantová teorie pole.[91]
Reference
- ^ Tiselius, A (1939). "Chemie proteinů a aminokyselin". Roční přehled biochemie. 8: 155–184. doi:10.1146 / annurev.bi.08.070139.001103.
- ^ A b Svedberg, T (1929). "Hmotnost a velikost proteinových molekul". Příroda. 123 (3110): 871. Bibcode:1929Natur.123..871S. doi:10.1038 / 123871a0. S2CID 4068088.
- ^ A b Svedberg, T (1934). "Sedimentace molekul v odstředivých polích". Chemické recenze. 14: 1–15. doi:10.1021 / cr60047a001.
- ^ A b C Bergmann, M; Niemann C (1937). „O struktuře bílkovin: hemoglobin skotu, vaječný albumin, fibrin skotu a želatina“. Journal of Biological Chemistry. 118: 301–314.
- ^ Svedberg, T (1930). "Oblasti stability pH proteinů". Transakce Faradayovy společnosti. 26: 741–744. doi:10.1039 / TF9302600737.
- ^ Fruton, JS (1979). "Rané teorie struktury proteinů". Annals of the New York Academy of Sciences. 325 (1): 1–18. Bibcode:1979NYASA.325 .... 1F. doi:10.1111 / j.1749-6632.1979.tb14125.x. PMID 378063. S2CID 39125170.
- ^ Hofmeister, F (1902). „Über Bau und Gruppierung der Eiweisskörper“. Ergebnisse der Physiologie. 1: 759–802. doi:10.1007 / BF02323641. S2CID 101988911.
- ^ Fischer, E. (1902). „Über die Hydrolyse der Proteinstoffe“. Chemiker Zeitung. 26: 939–940.
- ^ Fischer, E. (1913). „Synthese von Depsiden, Flechtenstoffen und Gerbstoffen“. Berichte der Deutschen Chemischen Gesellschaft. 46 (3): 3253–3289. doi:10.1002 / cber.191304603109.
- ^ Sørensen, SPL (1930). "Konstituce rozpustných proteinů jako reverzibilně disociovatelných komponentních systémů". Comptes Rendus des Travaux du Laboratoire Carlsberg. 18: 1–124.
- ^ Fruton, JS (1999). Proteiny, enzymy, geny: souhra chemie a biologie. New Haven, CT: Yale University Press. ISBN 0-585-35980-6.
- ^ Abderhalden, E. (1924). "Diketopiperaziny". Naturwissenschaften. 12 (36): 716–720. Bibcode:1924NW ..... 12..716A. doi:10.1007 / BF01504819. S2CID 29012795.
- ^ Abderhalden, E.; Komm E (1924). „Über die Anhydridstruktur der Proteine“. Zeitschrift für Physiologische Chemie. 139 (3–4): 181–204. doi:10.1515 / bchm2.1924.139.3-4.181.
- ^ Linderstrøm-Lang, K; Hotchkiss RD; Johansen G (1938). "Peptidové vazby v globulárních proteinech". Příroda. 142 (3605): 996. Bibcode:1938Natur.142..996L. doi:10.1038 / 142996a0. S2CID 4086716.
- ^ Chick, H; Martin CJ (1910). "Na" tepelnou "koagulaci proteinů". Fyziologický časopis. 40 (5): 404–430. doi:10.1113 / jphysiol.1910.sp001378. PMC 1533708. PMID 16993016.
Chick, H; Martin CJ (1911). „Na„ tepelnou “koagulaci proteinů. II. Působení horké vody na vaječný albumin a vliv kyselin a solí na rychlost reakce“. Fyziologický časopis. 43 (1): 1–27. doi:10.1113 / jphysiol.1911.sp001456. PMC 1512746. PMID 16993081.
Chick, H; Martin CJ (1912). „Na„ tepelnou “koagulaci proteinů. III. Vliv zásady na rychlost reakce“. Fyziologický časopis. 45 (1–2): 61–69. doi:10.1113 / jphysiol.1912.sp001535. PMC 1512881. PMID 16993182.
Chick, H; Martin CJ (1912). „Na„ tepelnou “koagulaci proteinů. IV. Podmínky regulující aglutinaci proteinů, na které již působila horká voda“. Fyziologický časopis. 45 (4): 261–295. doi:10.1113 / jphysiol.1912.sp001551. PMC 1512885. PMID 16993156. - ^ Anson, ML; Mirsky AE (1929). „Srážení bílkovin a jejich zvrácení“. Journal of General Physiology. 13 (2): 121–132. doi:10.1085 / jgp.13.2.121. PMC 2141032. PMID 19872511.
- ^ A b Anson, ML (1945). "Denaturace proteinů a vlastnosti proteinových skupin". Pokroky v chemii proteinů. 2: 361–386. doi:10.1016 / S0065-3233 (08) 60629-4. ISBN 978-0-12-034202-0.
- ^ Wu, H (1931). „Studie o denaturaci proteinů. XIII. Teorie denaturace“. Čínský žurnál fyziologie. 5: 321–344. Předběžné zprávy byly předloženy před XIII. Mezinárodním kongresem fyziologie v Bostonu (19. – 24. Srpna 1929) a v říjnovém čísle 1929 American Journal of Physiology.
- ^ A b Mirsky, AE; Pauling L. (1936). „O struktuře nativních, denaturovaných a koagulovaných proteinů“ (PDF). Sborník Národní akademie věd Spojených států amerických. 22 (7): 439–447. Bibcode:1936PNAS ... 22..439M. doi:10.1073 / pnas.22.7.439. PMC 1076802. PMID 16577722.
- ^ Neurath, H; Greenstein JP; Putnam FW; Erickson JO (1944). "Chemie denaturace proteinů". Chemické recenze. 34 (2): 157–265. doi:10.1021 / cr60108a003.
- ^ Putnam, F (1953). "Denaturace bílkovin". Proteiny (H. Neurath a K. Bailey, ed.). 1B: 807–892.
- ^ Astbury, WT; Woods HJ (1931). "Molekulová hmotnost proteinů". Příroda. 127 (3209): 663–665. Bibcode:1931Natur.127..663A. doi:10.1038 / 127663b0. S2CID 4133226.
- ^ Astbury, WT (1933). „Některé problémy v rentgenové analýze struktury zvířecích chloupků a jiných bílkovinných vláken“. Transakce Faradayovy společnosti. 29 (140): 193–211. doi:10.1039 / tf9332900193.
- ^ A b Jordan Lloyd, D (1932). "Koloidní struktura a její biologický význam". Biologické recenze. 7 (3): 254–273. doi:10.1111 / j.1469-185x.1962.tb01043.x. S2CID 84286671.
- ^ Jordan Lloyd, D; Marriott (1933). Msgstr "Neznámý název". Transakce Faradayovy společnosti. 29: 1228. doi:10.1039 / tf9332901228.
- ^ Astbury, WT (1936). "Neznámý název". Journal of the Textile Institute. 27: 282–?.
- ^ Wrinch, DM (1936). "Vzorec bílkovin". Příroda. 137 (3462): 411–412. Bibcode:1936Natur.137..411W. doi:10.1038 / 137411a0. S2CID 4140591.
- ^ Wrinch, DM (1936). "Energie formování molekul 'Cyclol'". Příroda. 138 (3484): 241–242. Bibcode:1936Natur.138..241W. doi:10.1038 / 138241a0. S2CID 4103892.
- ^ Frank, FC (1936). "Energie formování molekul 'Cyclol'". Příroda. 138 (3484): 242. Bibcode:1936Natur.138..242F. doi:10.1038 / 138242a0. S2CID 4065283.
- ^ Langmuir, já; Wrinch DM (1939). "Povaha cyklického svazku". Příroda. 143 (3611): 49–52. Bibcode:1939Natur.143 ... 49L. doi:10.1038 / 143049a0. S2CID 4056966.
- ^ Langmuir, já (1939). "Struktura proteinů". Sborník Fyzikální společnosti. 51 (4): 592–612. Bibcode:1939PPS .... 51..592L. doi:10.1088/0959-5309/51/4/305.
- ^ Wrinch, DM (1938). „O hydrataci a denaturaci proteinů“. Filozofický časopis. 25: 705–739.
- ^ Wrinch, DM (1936). "Hydratace a denaturace proteinů". Příroda. 142 (3588): 260. Bibcode:1938Natur.142..259.. doi:10.1038 / 142259a0.
- ^ Dow, RB; Matthews JE; Jr. and Thorp WTS (1940). "Účinek vysokotlakého ošetření na fyziologickou aktivitu inzulínu". American Journal of Physiology. 131 (2): 382–387. doi:10.1152 / ajplegacy.1940.131.2.382.
- ^ Kauzmann, W (1959). "Některé faktory při interpretaci denaturace proteinů". Pokroky v chemii proteinů. 14: 1–63. doi:10.1016 / S0065-3233 (08) 60608-7. ISBN 978-0-12-034214-3. PMID 14404936.
- ^ Wrinch, DM (1936). "Struktura proteinů a určitých fyziologicky aktivních sloučenin". Příroda. 138 (3493): 651–652. Bibcode:1936Natur.138..651W. doi:10.1038 / 138651a0. S2CID 4108696.
- ^ Wrinch, DM; Jordan Lloyd D (1936). „Vodíková vazba a struktura proteinů“. Příroda. 138 (3496): 758–759. Bibcode:1936Natur.138..758W. doi:10.1038 / 138758a0. S2CID 4096438.
- ^ Wrinch, DM (1937). "Povaha vazby v proteinech". Příroda. 139 (3521): 718. Bibcode:1937Natur.139..718W. doi:10.1038 / 139718a0. S2CID 4116000.
- ^ Astbury, WT; Wrinch DM (1937). "Intramolekulární skládání proteinů pomocí keto-enolové výměny". Příroda. 139 (3523): 798. Bibcode:1937Natur.139..798A. doi:10.1038 / 139798a0. S2CID 41311699.
- ^ Wrinch, D. "Fabric Theory of Protein Structure". Filozofický časopis. 30: 64–67.
- ^ Wrinch, DM (1937). „Cyklická teorie a„ globulární “proteiny“. Příroda. 139 (3527): 972–973. Bibcode:1937Natur.139..972W. doi:10.1038 / 139972a0. S2CID 4066210.
- ^ Wrinch, DM (1947). "Nativní protein". Věda. 106 (2743): 73–76. Bibcode:1947Sci ... 106 ... 73W. doi:10.1126 / science.106.2743.73. PMID 17808858.
- ^ Wrinch, DM (1937). "Na vzoru bílkovin". Sborník Královské společnosti. A160: 59–86.
Wrinch, DM (1937). „Cyklická hypotéza a„ globulární “proteiny. Sborník Královské společnosti. A161: 505–524.
Wrinch, DM (1938). „O molekulárních hmotnostech globulárních proteinů“. Filozofický časopis. 26: 313–332. - ^ Wrinch, DM (1939). "Tuberculin Protein TBU-skot (523)". Příroda. 144 (3636): 77. Bibcode:1939Natur.144 ... 77W. doi:10.1038 / 144077a0. S2CID 4113033.
- ^ Wrinch, DM (1937). "Na strukturu pepsinu". Filozofický časopis. 24: 940.
- ^ Wrinch, DM (1938). "Struktura pepsinu". Příroda. 142 (3587): 217. Bibcode:1938Natur.142..215.. doi:10.1038 / 142215a0.
- ^ Wrinch, DM (1937). „O struktuře inzulínu“. Věda. 85 (2215): 566–567. Bibcode:1937Sci .... 85..566W. doi:10.1126 / science.85.2215.566. PMID 17769864.
Wrinch, DM (1937). „O struktuře inzulínu“. Transakce Faradayovy společnosti. 33: 1368–1380. doi:10.1039 / tf9373301368.
Wrinch, DM (1938). "Struktura molekuly inzulínu". Journal of the American Chemical Society. 60 (8): 2005–2006. doi:10.1021 / ja01275a514.
Wrinch, DM (1938). "Struktura molekuly inzulínu". Věda. 88 (2276): 148–149. Bibcode:1938Sci .... 88..148W. doi:10.1126 / science.88.2276.148-a. PMID 17751525.
Wrinch, DM; Langmuir I. (1938). "Struktura molekuly inzulínu". Journal of the American Chemical Society. 60 (9): 2247–2255. doi:10.1021 / ja01276a062.
Langmuir, já; Wrinch DM (1939). "Poznámka ke struktuře inzulínu". Sborník Fyzikální společnosti. 51 (4): 613–624. Bibcode:1939PPS .... 51..613L. doi:10.1088/0959-5309/51/4/306. - ^ Bragg, WL (1939). "Pattersonovy diagramy v krystalové analýze". Příroda. 143 (3611): 73–74. Bibcode:1939Natur.143 ... 73B. doi:10.1038 / 143073a0. S2CID 4063833.
Bernal, JD (1939). "Vektorové mapy a cyklická hypotéza". Příroda. 143 (3611): 74–75. Bibcode:1939Natur.143 ... 74B. doi:10.1038 / 143074a0. S2CID 4108005.
Robertson, JM (1939). "Vektorové mapy a těžké atomy v krystalové analýze a struktuře inzulínu". Příroda. 143 (3611): 75–76. Bibcode:1939Natur.143 ... 75R. doi:10.1038 / 143075a0. S2CID 4053119. - ^ Riley, DP; Fankuchen I (1939). „Odvozená Pattersonova analýza kostry Cyclolu C.2 Molekula". Příroda. 143 (3624): 648–649. Bibcode:1939Natur.143..648R. doi:10.1038 / 143648a0. S2CID 4086672.
Wrinch, DM (1940). „Pattersonova projekce koster struktury navržená pro molekulu inzulínu“. Příroda. 145 (3687): 1018. Bibcode:1940Natur.145.1018W. doi:10.1038 / 1451018a0. S2CID 4119581.
Riley, D (1940). „Pattersonova analýza odvozená z Cyclol C.2 Skeleton". Příroda. 146 (3694): 231. Bibcode:1940Natur.146..231R. doi:10.1038/146231a0. S2CID 4111977. - ^ Neurath, H; Bull HB (1938). "The Surface Activity of Proteins". Chemické recenze. 23 (3): 391–435. doi:10.1021/cr60076a001.
- ^ Huggins, M (1939). "The Structure of Proteins". Journal of the American Chemical Society. 61 (3): 755. doi:10.1021/ja01872a512.
- ^ Haurowitz, F (1938). "Die Anordnung der Peptidketten in Sphäroprotein-Molekülen". Hoppe-Seyler's Zeitschrift für Physiologische Chemie. 256: 28–32. doi:10.1515/bchm2.1938.256.1.28.
- ^ Meyer, KH; Hohenemser W (1938). "Possibility of the Formation of Cyclols from Simple Peptides". Příroda. 141 (3582): 1138–1139. Bibcode:1938Natur.141.1138M. doi:10.1038/1411138b0. S2CID 4097115.
- ^ Bergmann, M; Niemann C (1938). "The Chemistry of Amino Acids and Proteins". Roční přehled biochemie. 7 (2): 99–124. doi:10.1146/annurev.bi.07.070138.000531. PMC 537431.
- ^ Neuberger, A (1939). "Chemical criticism of the cyclol and frequency hypothesis of protein structure". Sborník Královské společnosti. 170: 64–65.
- ^ Neuberger, A (1939). "Chemical Aspects of the Cyclol Hypothesis". Příroda. 143 (3620): 473. Bibcode:1939Natur.143..473N. doi:10.1038/143473a0. S2CID 4102966.
- ^ Haurowitz, F; Astrup T (1939). "Ultraviolet absorption of genuine and hydrolysed protein". Příroda. 143 (3612): 118–119. Bibcode:1939Natur.143..118H. doi:10.1038/143118b0. S2CID 4078416.
- ^ Klotz, IM; Griswold P (1949). "Infrared Spectra and the Amide Linkage in a Native Globular Protein". Věda. 109 (2830): 309–310. Bibcode:1949Sci...109..309K. doi:10.1126/science.109.2830.309. PMID 17782718.
- ^ A b Pauling, L; Niemann C (1939). "The Structure of Proteins". Journal of the American Chemical Society. 61 (7): 1860–1867. doi:10.1021/ja01876a065.
- ^ Hotchkiss, RD (1939). "The Determination of Peptide Bonds in Crystalline Lactoglobulin". Journal of Biological Chemistry. 131: 387–395.
- ^ A b Wrinch, DM (1941). "The Geometrical Attack on Protein Structure". Journal of the American Chemical Society. 63 (2): 330–33. doi:10.1021/ja01847a004.
- ^ Wrinch, DM (1940). "The Cyclol Hypothesis". Příroda. 145 (3678): 669–670. Bibcode:1940Natur.145..669W. doi:10.1038/145669a0. S2CID 4065545.
- ^ A b Wrinch, DM (1948). "The Native Proteins as Polycondensations of Amino Acids". Věda. 107 (2783): 445–446. Bibcode:1948Sci...107R.445W. doi:10.1126/science.107.2783.445-a. PMID 17844448.
- ^ Wrinch, DM (1948). "Skeletal Units in Protein Crystals". Věda. 115 (2987): 356–357. Bibcode:1952Sci...115..356W. doi:10.1126/science.115.2987.356. PMID 17748855.
- ^ Wrinch, DM (1948). "Molecules of the Insulin Structure". Věda. 116 (3021): 562–564. Bibcode:1952Sci...116..562W. doi:10.1126/science.116.3021.562. PMID 13015111.
- ^ Wrinch, DM (1939). "The Structure of the Globular Proteins". Příroda. 143 (3620): 482–483. Bibcode:1939Natur.143..482W. doi:10.1038/143482a0. S2CID 5362977.
- ^ Wrinch, DM (1939). "The Cyclol Theory and the Structure of Insulin". Příroda. 143 (3627): 763–764. Bibcode:1939Natur.143..763W. doi:10.1038/143763a0. S2CID 4063795.
- ^ Wrinch, DM (1939). "Native Proteins, Flexible Frameworks and Cytoplasmic Organization". Příroda. 150 (3800): 270–271. Bibcode:1942Natur.150..270W. doi:10.1038/150270a0. S2CID 4085657.
- ^ Anslow, GA (1942). "Bond Energies in Some Protein Fabrics and Side Chains". Fyzický přehled. 61 (7–8): 547. Bibcode:1942PhRv...61..541.. doi:10.1103/PhysRev.61.541.
- ^ Anslow, GA (1945). "Ultraviolet Spectra of Biologically Important Molecules". Journal of Applied Physics. 16 (1): 41–49. Bibcode:1945JAP....16...41A. doi:10.1063/1.1707499.
- ^ Anslow, GA (1953). "The Sites of the Amino-Acid Residues on a Cyclol Model of Insulin". Journal of Chemical Physics. 21 (11): 2083–2084. Bibcode:1953JChPh..21.2083A. doi:10.1063/1.1698765.
- ^ Guedez, T; Núñez A; Tineo E; Núñez O (2002). "Ring size configuration effect and the transannular intrinsic rates in bislactam macrocycles". Journal of the Chemical Society, Perkin Transactions 2. 2002 (12): 2078–2082. doi:10.1039/b207233e.
- ^ Bernal, JD (1939). "Structure of proteins". Příroda. 143 (3625): 663–667. Bibcode:1939Natur.143..663B. doi:10.1038/143663a0. S2CID 46327591.
- ^ Wieland T and Bodanszky M, The World of Peptides, Springer Verlag, pp.193–198. ISBN 0-387-52830-X
- ^ Hofmann, A; Ott H; Griot R; Stadler PA; Frey AJ (1963). "Synthese von Ergotamin". Helvetica Chimica Acta. 46: 2306–2336. doi:10.1002/hlca.19630460650.
- ^ Shemyakin, MM; Antonov VK; Shkrob AM (1963). "Activation of the amide group by acylation". Peptides, Proc. 6th Europ. Pept. Symp., Athens: 319–328.
- ^ Zanotti, G; Pinnen F; Lucente G; Cerrini S; Fedeli W; Mazza F (1984). "Peptide thiacyclols. Synthesis and structural studies". J. Chem. Soc. Perkin Trans. 1: 1153–1157. doi:10.1039/p19840001153.
- ^ Griot, RG; Frey AJ (1963). "The formation of cyclols from N-hydroxyacyl lactames". Čtyřstěn. 19 (11): 1661–1673. doi:10.1016/S0040-4020(01)99239-7.
- ^ Lucente, G; Romeo A (1971). "Synthesis of cyclols from small peptides via amide-amide reaction". Chem. Commun. ?: 1605–1607. doi:10.1039/c29710001605.
Rothe M, Schindler W, Pudill R, Kostrzewa U, Theyson R, and Steinberger R. (1971) "Zum Problem der Cycloltripeptidsynthese", Peptides, Proc. 11th Europ. Pept. Symp., Wien, 388–399.
Rothe M and Roser KL. (1988) "Conformational flexibility of cyclic tripeptides", Abstr. 20th Europ. Pept. Symp. Tübingen, str. 36. - ^ Wieland T and Mohr H. (1956) "Diacylamide als energiereiche Verbindungen. Diglycylimid", Liebigs Ann. Chem., 599, 222–232.
Wieland T and Urbach H. (1958) "Weitere Di-Aminoacylimide und ihre intramolekulare Umlagerung", Liebigs Ann. Chem., 613, 84–95.
Brenner, M (1958). Wolstenholme GEW; O'Connor CM (eds.). "The aminoacyl insertion". Ciba Foundation Symposium on Amino Acids and Peptides with Antimetabolic Activity. - ^ Wrinch, DM (1957). "Structure of Bacitracin A". Příroda. 179 (4558): 536–537. Bibcode:1957Natur.179..536W. doi:10.1038/179536a0. S2CID 4154444.
Wrinch, DM (1957). "An Approach to the Synthesis of Polycyclic Peptides". Příroda. 180 (4584): 502–503. Bibcode:1957Natur.180..502W. doi:10.1038/180502b0. S2CID 4289278.
Wrinch, DM (1962). "Some Issues in Molecular Biology and Recent Advances in the Organic Chemistry of Small Peptides". Příroda. 193 (4812): 245–247. Bibcode:1962Natur.193..245W. doi:10.1038/193245a0. PMID 14008494. S2CID 4252124.
Wrinch, DM (1963). "Recent Advances in Cyclol Chemistry". Příroda. 199 (4893): 564–566. Bibcode:1963Natur.199..564W. doi:10.1038/199564a0. S2CID 4177095.
Wrinch, DM (1965). "A Contemporary Picture of the Chemical Aspects of Polypeptide Chain Structures and Certain Problems of Molecular Biology". Příroda. 206 (4983): 459–461. Bibcode:1965Natur.206..459W. doi:10.1038/206459a0. PMID 5319104. S2CID 4190760. - ^ Wrinch, DM (1960). Chemical Aspects of the Structures of Small Peptides: An Introduction. Kodaň: Munksgaard.
- ^ Wrinch, DM (1965). Chemical Aspects of Polypeptide Chain Structures and the Cyclol Theory. New York: Plenum Press.
- ^ Kauzmann, W (1993). "Reminiscences from a life in protein physical chemistry". Věda o bílkovinách. 2 (4): 671–691. doi:10.1002/pro.5560020418. PMC 2142355. PMID 8518739.
- ^ Tanford, C; Reynolds J (2001). Nature's robots: A history of proteins. Oxford: Oxford University Press. ISBN 0-19-850466-7.
- ^ Pais, A (1986). Inward Bound: of Matter and Forces in the Physical World. Oxford University Press. ISBN 0-19-851971-0.
Bohr, N (1913). "On the Constitution of Atoms and Molecules (Part 1 of 3)". Filozofický časopis. 26: 1–25. Bibcode:1913PMag...26....1B. doi:10.1080/14786441308634955. Archivovány od originál on 2007-07-04.
Bohr, N (1913). "On the Constitution of Atoms and Molecules, Part II Systems Containing Only a Single Nucleus". Filozofický časopis. 26 (153): 476–502. Bibcode:1913PMag ... 26..476B. doi:10.1080/14786441308634993.
Bohr, N (1913). "On the Constitution of Atoms and Molecules, Part III". Filozofický časopis. 26: 857–875. Bibcode:1913PMag...26..857B. doi:10.1080/14786441308635031.
Bohr, N (1914). "The spectra of helium and hydrogen". Příroda. 92 (2295): 231–232. Bibcode:1913Natur..92..231B. doi:10.1038/092231d0. S2CID 11988018. - ^ Pauling, L; Corey RB (1953). "A Proposed Structure For The Nucleic Acids". Sborník Národní akademie věd. 39 (2): 84–97. Bibcode:1953PNAS ... 39 ... 84P. doi:10.1073/pnas.39.2.84. PMC 1063734. PMID 16578429.
- ^ Franklin, RE; Gosling R (1953). "Molecular configuration of sodium thymonucleate". Příroda. 171 (4356): 740–741. Bibcode:1953Natur.171..740F. doi:10.1038/171740a0. PMID 13054694. S2CID 4268222.
- ^ A b Watson, JD; Crick F (1953). "Molecular structure of nucleic acids: A structure for deoxyribonucleic acid". Příroda. 171 (4356): 737–738. Bibcode:1953Natur.171..737W. doi:10.1038 / 171737a0. PMID 13054692. S2CID 4253007.
- ^ Saenger, W (1988). Principles of Nucleic Acid Structure. Springer Verlag. ISBN 0-387-90762-9.
- ^ Pais, A. (1982). Subtle is the Lord: The Science and the Life of Albert Einstein. Oxford University Press. ISBN 0-19-853907-X.
Další čtení
- "Protein Units Put in Graphic 'Cage'", The New York Times, str. 14, 19 April 1940.
- "Waffle-Iron Theory of Proteins", The New York Times, str. E9, 2 February 1947.
- Senechal, Marjorie, vyd. (1980), Structures of Matter and Patterns in Science, inspired by the work and life of Dorothy Wrinch, 1894–1976: Proceedings of a Symposium held at Smith College, Northampton, Massachusetts on 28–30 September 1977, Schenkman Publishing Company.
- "Selected papers of Dorothy Wrinch, from the Sophia Smith Collection", in Structures of Matter and Patterns in Science.
- Senechal, Marjorie (2013), I Died For Beauty: Dorothy Wrinch and the Cultures of Science, Oxford University Press.