Hnědé řasy - Brown algae
| Hnědé řasy | |
|---|---|
 | |
| Obří řasa (Macrocystis pyrifera ) | |
| Vědecká klasifikace | |
| Clade: | SAR |
| Kmen: | Ochrophyta |
| Podkmen: | Phaeista |
| Infraphylum: | Limnista |
| Nadtřída: | Fucistia |
| Třída: | Phaeophyceae Kjellman, 1891[3] |
| Objednávky | |
Vidět klasifikace | |
| Synonyma | |
| |
The hnědé řasy (singulární: řasa), zahrnující třída Phaeophyceae, jsou velkou skupinou mnohobuňečný řasy, včetně mnoha mořských řas umístěných v chladnějších vodách v rámci EU Severní polokoule. Většina hnědých řas žije v mořském prostředí, kde hrají důležitou roli jako potrava i jako potenciál místo výskytu. Například, Makrocystis, a kelp objednávky Laminariales, může dosáhnout délky 60 m (200 ft) a tvoří prominentní pod vodou řasy. Řasy podobné těmto lesům obsahují vysokou úroveň biologické rozmanitosti.[4] Dalším příkladem je Sargassum, který vytváří jedinečné plovoucí rohože z mořských řas v tropických vodách Sargasové moře které slouží jako stanoviště mnoha druhů. Mnoho hnědých řas, například členové řádu Fucales, běžně rostou podél skalnatých mořských pobřeží. Někteří členové třídy, například řasy, lidé používají jako jídlo.
Mezi 1 500 a 2 000 druh hnědých řas je známo po celém světě.[5] Některé druhy, jako např Ascophyllum nodosum, se kvůli své obchodní důležitosti stali samostatnými subjekty rozsáhlého výzkumu. Mají také environmentální význam uhlíková fixace.[4]
Hnědé řasy patří do skupiny Heterokontophyta, velká skupina eukaryotický organismy se vyznačují nejvýrazněji tím, že mají chloroplasty obklopen čtyřmi membránami, což naznačuje původ z a symbiotický vztah mezi bazálním eukaryotem a jiným eukaryotickým organismem. Většina hnědých řas obsahuje pigment fucoxanthin, který je zodpovědný za výraznou zelenohnědou barvu, která jim dává jejich jméno. Hnědé řasy jsou mezi heterokonty jedinečné ve vývoji do mnohobuněčných forem s diferencované papírové kapesníky, ale reprodukují se pomocí bičované výtrusy a gamety které se velmi podobají buňkám jiných heterokontů. Genetické studie ukazují, že jejich nejbližší příbuzní jsou žlutozelené řasy.
Morfologie
Hnědé řasy existují v široké škále velikostí a forem. Nejmenší členové skupiny rostou jako drobné, nadýchané chomáčky nitkovitých vláken buňky ne delší než několik centimetrů.[6] Některé druhy mají ve svém životním cyklu fázi, která se skládá pouze z několika buněk, takže celá řasa je mikroskopická. Jiné skupiny hnědých řas dorůstají do mnohem větších velikostí. The skalničky a kožovité řasy jsou často nejnápadnějšími řasami na jejich stanovištích.[7] Kelpy mohou mít velikost od 60 centimetrů vysoké mořské palmy Postelsia k obřím řasám Macrocystis pyrifera, který roste na více než 50 m (150 stop) dlouhý[8][9] a je největší ze všech řas. Ve formě se hnědé řasy pohybují od malých krust nebo polštářů[10] na listnaté volně plovoucí podložky tvořené druhy Sargassum. Mohou se skládat z jemných plstnatých pramenů buněk, jako v Ectocarpus, nebo 30 centimetrů dlouhé (1 stopy) zploštělé větve připomínající ventilátor, jako v Padina.
Bez ohledu na velikost nebo formu oddělují Phaeophyceae dva viditelné rysy od všech ostatních řas. Za prvé, členové skupiny mají charakteristickou barvu, která se pohybuje od olivově zelená do různých odstínů hnědý. Konkrétní odstín závisí na množství fucoxanthin přítomný v řase.[11] Zadruhé, všechny hnědé řasy jsou mnohobuňečný. Nejsou známy žádné druhy, které existují jako jednotlivé buňky nebo jako kolonie buněk,[11] a hnědé řasy jsou jedinou hlavní skupinou mořské řasy který takové formy nezahrnuje. Může to však být spíše výsledkem klasifikace než důsledkem evoluce, protože všechny skupiny předpokládané jako nejbližší příbuzní hnědých zahrnují jednobuněčné nebo koloniální formy. Mohou měnit barvu v závislosti na slanosti, od načervenalé po hnědou.
Viditelné struktury

Tělo všech hnědých řas je bez ohledu na jejich formu nazýváno a thallus, což naznačuje, že mu chybí komplex xylem a phloem z cévnatých rostlin. To neznamená, že hnědým řasám zcela chybí specializované struktury. Protože však někteří botanici definují „pravé“ stonky, listy a kořeny přítomností těchto tkání, jejich nepřítomnost v hnědých řasách znamená, že stonkové a listové struktury nalezené v některých skupinách hnědých řas musí být popsány pomocí jiná terminologie.[12] I když ne všechny hnědé řasy jsou strukturálně složité, ty, které mají obvykle jednu nebo více charakteristických částí.
A sevření je kořenová struktura přítomná ve spodní části řasy. Stejně jako kořenový systém v rostlinách slouží pevná kotva k ukotvení řasy na místě na Podklad kde roste, a tak brání tomu, aby řasa byla unesena proudem. Na rozdíl od kořenového systému zadržování obecně neslouží jako primární orgán pro příjem vody ani nepřijímá živiny ze substrátu. Celkový fyzický vzhled fixace se liší mezi různými hnědými řasami a mezi různými substráty. Může být silně rozvětvený nebo může vypadat jako kalich. Jedna řasa má obvykle jen jednu pevnost, i když u některých druhů roste z jejich pevnosti více než jedna třeně.
A stipe je stonková nebo stonková struktura přítomná v řase. Může růst jako krátká struktura blízko základny řasy (jako v Laminaria ), nebo se může vyvinout do velké složité struktury probíhající v celém těle řas (jako v Sargassum nebo Makrocystis ). V nejvíce strukturálně diferencovaných hnědých řasách (např Fucus ), tkáně uvnitř třeně jsou rozděleny do tří odlišných vrstev nebo oblastí. Mezi tyto oblasti patří centrální dřeň, okolní kůra a vnější epidermis, z nichž každá má analog ve stonku cévnaté rostliny. U některých hnědých řas zahrnuje dřeňová oblast jádro podlouhlých buněk, které se podobají phloem cévnatých rostlin jak ve struktuře, tak ve funkci. V ostatních (např Nereocystis ), střed třeně je dutý a naplněný plynem, který slouží k udržení této části řas vznášející se. Řapík může být u druhů relativně pružný a pružný Macrocystis pyrifera které rostou v silných proudech, nebo mohou být u druhů podobných přísnější Postelsia palmaeformis které jsou vystaveny atmosféře při odlivu.
Mnoho řas má zploštělou část, která může připomínat list, a to se nazývá a čepel, plátek nebo vějířovitý. Název čepel se nejčastěji aplikuje na jednu nerozdělenou strukturu, zatímco vějířovitý lze použít na všechny nebo většinu řas, které jsou zploštělé, ale tento rozdíl se všeobecně nepoužívá. Název plátek Výraz "strukturálně diferencovaná řasa" označuje část strukturálně diferencované řasy, která je zploštělá. Může to být jednoduchá nebo rozdělená struktura a může být rozprostřena po podstatné části řasy. v skalničky například lamina je široké křídlo tkáně, které běží nepřetržitě po obou stranách rozvětveného midrib. Střední část a lamina společně tvoří téměř všechny skalní řasy, takže se lamina šíří po celé řase, spíše než existuje jako její lokalizovaná část.

V některých hnědých řasách je jedna vrstva nebo čepel, zatímco v jiných může být mnoho samostatných čepelí. Dokonce i u těch druhů, které zpočátku produkují jedinou čepel, se struktura může roztrhat drsnými proudy nebo jako součást zrání za vzniku dalších čepelí. Tyto čepele mohou být připevněny přímo k třeně, k upevnění bez přítomnosti třeně, nebo může být mezi třmenem a čepelí vzduchový měchýř. Povrch laminy nebo čepele může být hladký nebo zvrásněný; její tkáně mohou být tenké a pružné nebo silné a kožovité. U druhů podobných Egregia menziesii, tato vlastnost se může měnit v závislosti na turbulencích vod, ve kterých roste.[6] U jiných druhů je povrch čepele potažen slizem, aby se zabránilo připojení epifity nebo odradit býložravci. Čepele jsou také často částmi řas, které nesou reprodukční struktury.
Volaly plynové plováky pneumatocyst poskytnout vztlak v mnoha řasy a členové Fucales. Tyto struktury podobné močovému měchýři se vyskytují v nebo v blízkosti plátek, takže se drží blíže k vodní hladině a přijímá tak více světla pro fotosyntézu. Pneumatocysty jsou nejčastěji sférické nebo elipsoidní, ale může se lišit tvarem u různých druhů. Druhy jako např Nereocystis luetkeana a Pelagophycus porra nést jednu velkou pneumatocystu mezi horní částí třeně a základnou lopatek. Naproti tomu obří řasa Macrocystis pyrifera nese mnoho čepelí podél jeho třeně, s pneumatocystou na základně každé čepele, kde se připojuje k hlavnímu třeně. Druhy Sargassum také nést mnoho lopatek a pneumatocyst, ale oba druhy struktur jsou připevněny samostatně k třeně krátkými stopkami. U druhů Fucus se pneumatocysty vyvíjejí uvnitř samotné laminy, buď jako diskrétní sférické měchýře, nebo jako podlouhlé oblasti naplněné plynem, které zaujímají obrys laminy, ve které se vyvíjejí.
Růst

K hnědým řasám patří největší a nejrychleji rostoucí mořské řasy.[6] Frondy z Makrocystis může dorůst až 50 cm za den a čípky mohou za jediný den dorůst o 6 cm.[13]
K růstu většiny hnědých řas dochází na špičkách struktur v důsledku rozdělení v jedné apikální buňka nebo v řadě takových buněk. Jsou to jednotlivé buněčné organismy.[7] Jak se tato apikální buňka dělí, nové buňky, které produkuje, se vyvíjejí do všech tkání řasy. Větvení a další boční struktury se objeví, když se apikální buňka rozdělí a vytvoří dvě nové apikální buňky. Několik skupin (např Ectocarpus ) rostou difúzní, nelokalizovanou produkcí nových buněk, které se mohou vyskytovat kdekoli na holi.[11]
Organizace tkání
Nejjednodušší hnědé jsou vláknité - to znamená, že jejich buňky jsou podlouhlé a mají přes svou šířku řezy septa. Rozvětvují se tím, že se na špičce rozšiřují a poté rozdělí rozšíření.[14]
Tato vlákna mohou být haplostichózní nebo polystichiální, multiaxiální nebo monoaxiální formující nebo ne pseudoparenchyma.[15][16] Kromě lístků existují i velké parenchyma tic kelps s trojrozměrným vývojem a růstem a různými tkáněmi (meristoderm, kůra a dřeň ), které lze považovat za mořské stromy.[17][18] K dispozici jsou také Fucales a Dictyotales menší než řasy, ale stále parenchymatický se stejným druhem odlišných tkání.
Buněčná stěna se skládá ze dvou vrstev; vnitřní vrstva nese pevnost a sestává z celulózy; vrstva vnější stěny je hlavně alginová a je vlhká, když je mokrá, ale po vyschnutí se stává tvrdou a křehkou.[15] Konkrétně se hnědá buněčná stěna řas skládá z několika složek, přičemž hlavními složkami jsou algináty a supratovaný fukan, z nichž každá tvoří až 40%.[19] Celulóza, hlavní složka většiny buněčných stěn rostlin, je přítomna ve velmi malém procentu, až 8%. Zdá se, že dráhy biosyntézy celulózy a alginátu byly získány z jiných organismů endosymbiotickým a horizontálním přenosem genů, zatímco suphated polysacharidy jsou původem.[20] Konkrétně se zdá, že celulózové syntázy pocházejí z červeného řasového endosymbiontu předka fotosyntetických stramenopilů a ansestor hnědých řas získal klíčové enzymy pro biosyntézu alginátů z aktinobakteria. Přítomnost a jemná kontrola struktury alginátu v kombinaci s celulózou, která existovala před ní, dala potenciálně hnědým řasám schopnost vyvíjet složité strukturně mnohobuněčné organismy, jako jsou řasy.[21]
Evoluční historie
Genetické a ultrastrukturální důkazy řadí Phaeophyceae mezi heterokonty (Stramenopiles),[22] velké shromáždění organismů, které zahrnuje obojí fotosyntetický členové s plastidy (tak jako rozsivky ), stejně jako nefotosyntetické skupiny (například slizové sítě a vodní formy ). Ačkoli některým příbuzným heterokontů hnědých řas chybí plastidy ve svých buňkách, vědci se domnívají, že je to spíše výsledek evoluční ztráty této organely v těchto skupinách než nezávislé získání několika fotosyntetickými členy.[23] Předpokládá se tedy, že všechny heterokonty pocházejí z jediného heterotrofní předek, který se stal fotosyntetickým, když prošel plastidy endosymbióza jiné jednobuněčné eukaryoty.[24]
Nejbližší příbuzní hnědých řas zahrnují jednobuněčné a vláknité druhy, ale jednobuněčné druhy hnědých řas nejsou známy. Většina vědců však předpokládá, že se Phaeophyceae vyvinuly z jednobuněčných předků.[25] DNA porovnání sekvence také naznačuje, že hnědé řasy se vyvinuly z vláknitých Phaeothamniophyceae,[26] Xanthophyceae,[27] nebo Chrysophyceae[28] mezi 150[1] a před 200 miliony let.[2] V mnoha ohledech se vývoj hnědých řas podobá vývoji hnědých řas zelené řasy a červené řasy,[29] protože všechny tři skupiny mají složité mnohobuněčné druhy s střídání generací. Analýza 5S rRNA sekvence odhalují mnohem menší evoluční vzdálenosti mezi rody hnědých řas než mezi rody červených nebo zelených řas,[2][30] což naznačuje, že hnědé řasy se diverzifikovaly mnohem nověji než ostatní dvě skupiny.
Fosílie
Výskyt Phaeophyceae as fosilie je vzácný kvůli jejich obecně měkké povaze,[31] a vědci nadále diskutují o identifikaci některých nálezů.[32] Část problému s identifikací spočívá v konvergentní evoluce morfologií mezi mnoha hnědými a červenými řasami.[33] Většina fosilií řas z měkkých tkání si zachovává pouze zploštělý obrys, aniž by měly mikroskopické rysy, které umožňují spolehlivé rozlišení hlavních skupin mnohobuněčných řas. Mezi hnědými řasami pouze druhy rodu Padina ukládat významné množství minerálů do nebo kolem jejich buněčných stěn.[34] Jiné skupiny řas, jako je červené řasy a zelené řasy, mít řadu vápenatý členů. Z tohoto důvodu je pravděpodobnější, že zanechají důkazy ve fosilních záznamech než měkká těla většiny hnědých řas a častěji je lze přesně klasifikovat.[35]
Fosílie srovnatelné morfologicky s hnědými řasami jsou známy z vrstev starých jako svrchní Ordovik,[36] ale taxonomické afinita těchto fosílií dojmu není zdaleka jistá.[37] Tvrdí to dříve Ediacaran fosilie jsou hnědé řasy[38] od té doby byli propuštěni.[26] Zatímco mnoho uhlíkatý fosilie byly popsány z Precambrian, jsou obvykle zachovány jako zploštělé obrysy nebo fragmenty o délce pouze milimetrů.[39] Protože těmto fosíliím chybí diagnostické funkce pro identifikaci i na nejvyšší úrovni, jsou přiřazeny fosilním tvoří taxony podle jejich tvaru a dalších hrubých morfologických znaků.[40] Počet Devonský zkameněliny pojmenované fukoidy, od jejich podobnosti v obrysu s druhy v rodu Fucus, se ukázaly být spíše anorganické než skutečné fosilie.[31] Devonská megafosílie Prototaxity, který se skládá z masy vláken seskupených do kmenových os, byl považován za možnou hnědou řasu.[11] Moderní výzkum však upřednostňuje reinterpretaci této fosílie jako pozemské houba nebo houbovitý organismus.[41] Stejně tak fosilní Protosalvinie kdysi byla považována za možnou hnědou řasu, ale nyní je považována za ranou suchozemská rostlina.[42]
Počet Paleozoikum fosílie byly předběžně klasifikovány s hnědými řasami, ačkoli většina z nich byla také srovnávána se známými druhy červených řas. Phascolophyllaphycus má četné podlouhlé, nafouknuté čepele připojené k třeně. Jedná se o nejhojnější fosílie řas nalezené ve sbírce vyrobené z Karbon vrstvy v Illinois.[43] Každá dutá čepel nese až osm pneumatocyst na základně a zdá se, že tyčinky byly také duté a nafouknuté. Tato kombinace charakteristik je podobná určitým moderním rodům v pořadí Laminariales (řasy). Několik fosilií Drydenia a jediný exemplář Hungerfordia z horního devonu z New York byly také srovnávány s hnědými i červenými řasami.[33] Fosilie z Drydenia Skládají se z eliptické čepele připevněné k rozvětvené vláknité fixaci, na rozdíl od některých druhů Laminaria, Porphyra nebo Gigartina. Jediný známý exemplář Hungerfordia větví se dichotomicky do laloků a připomíná rody Chondrus a Fucus[33] nebo Dictyota.[44]
Nejdříve známé fosílie, které lze spolehlivě přiřadit k Phaeophyceae, pocházejí Miocén křemelina vklady Montereyské souvrství v Kalifornie.[24] Několik měkkých hnědých makrořas, jako např Julescraneia, byl nalezen.[45]
Klasifikace
Fylogeneze
Na základě práce Silberfelda, Rousseau & de Reviers 2014.[46]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Taxonomie
Toto je seznam objednávky ve třídě Phaeophyceae:[46][47]
- Třída Phaeophyceae Hansgirg 1886 [Fucophyceae; Melanophycidae Rabenhorst 1863 stat. listopad. Cavalier-Smith 2006]
- Podtřída Discosporangiophycidae Silberfeld, Rousseau & Reviers 2014
- Objednat Discosporangiales Schmidt 1937 emend. Kawai a kol. 2007
- Rodina Choristocarpaceae Kjellman 1891
- Rodina Discosporangiaceae Schmidt 1937
- Objednat Discosporangiales Schmidt 1937 emend. Kawai a kol. 2007
- Podtřída Ishigeophycidae Silberfeld, Rousseau & Reviers 2014
- Objednat Ishigeales Cho & Vypískat 2004
- Rodina Ishigeaceae Okamura 1935
- Rodina Petrodermataceae Silberfeld, Rousseau & Reviers 2014
- Objednat Ishigeales Cho & Vypískat 2004
- Podtřída Dictyotophycidae Silberfeld, Rousseau & Reviers 2014
- Objednat Dictyotales Bory de Saint-Vincent 1828 např Phillips et al.
- Rodina Dictyotaceae Lamouroux ex Dumortier 1822 [Scoresbyellaceae Womersley 1987; Dictyopsidaceae]
- Objednat Onslowiales Draisma & Prud’homme van Reine 2008
- Rodina Onslowiaceae Draisma & Prud’homme van Reine 2001
- Objednat Sphacelariales Migula 1909
- Rodina Cladostephaceae Oltmanns 1922
- Rodina Lithodermataceae Hauck 1883
- Rodina Phaeostrophiaceae Kawai a kol. 2005
- Rodina Sphacelariaceae Decaisne 1842
- Rodina Sphacelodermaceae Draisma, Prud’homme & Kawai 2010
- Rodina Stypocaulaceae Oltmanns 1922
- Objednat Syringodermatales Henry 1984
- Rodina Syringodermataceae Henry 1984
- Objednat Dictyotales Bory de Saint-Vincent 1828 např Phillips et al.
- Podtřída Fucophycidae Cavalier-Smith 1986
- Objednat Ascoseirales Petrov 1964 vychází. Moe a Henry 1982
- Rodina Ascoseiraceae Skottsberg 1907
- Objednat Asterocladales T. Silberfeld et al. 2011
- Rodina Asterocladaceae Silberfeld a kol. 2011
- Objednat Desmarestiales Setchell & Gardner 1925
- Rodina Arthrocladiaceae Chauvin 1842
- Rodina Desmarestiaceae (Thuret) Kjellman 1880
- Objednat Ectocarpales Bessey 1907 vychází. Rousseau & Reviers 1999a [Chordariales Setchell & Gardner 1925; Dictyosiphonales Setchell & Gardner 1925; Scytosiphonales Feldmann 1949]
- Rodina Acinetosporaceae Hamel ex Feldmann 1937 [Pylaiellaceae; Pilayellaceae]
- Rodina Adenocystaceae Rousseau a kol. 2000 emend. Silberfeld a kol. 2011 [Chordariopsidaceae]
- Rodina Chordariaceae Greville 1830 emend. Peters & Ramírez 2001 [Myrionemataceae]
- Rodina Ectocarpaceae Agardh 1828 emend. Silberfeld a kol. 2011
- Rodina Petrospongiaceae Racault a kol. 2009
- Rodina Scytosiphonaceae Ardissone & Straforello 1877 [Chnoosporaceae Setchell & Gardner 1925]
- Objednat Fucales Bory de Saint-Vincent 1827 [Notheiales Womersley 1987; Durvillaeales Petrov 1965]
- Rodina Bifurcariopsidaceae Cho a kol. 2006
- Rodina Durvillaeaceae (Oltmanns) De Toni 1891
- Rodina Fucaceae Adanson 1763
- Rodina Himanthaliaceae (Kjellman) De Toni 1891
- Rodina Hormosiraceae Fritsch 1945
- Rodina Notheiaceae Schmidt 1938
- Rodina Sargassaceae Kützing 1843 [Cystoseiraceae De Toni 1891]
- Rodina Seirococcaceae Nizamuddin 1987
- Rodina Xiphophoraceae Cho a kol. 2006
- Objednat Laminariales Migula 1909 [Phaeosiphoniellales Silberfeld, Rousseau & Reviers 2014 ord. listopad. podpěra.]
- Rodina Agaraceae Postels & Ruprecht 1840 [Costariaceae]
- Rodina Akkesiphycaceae Kawai & Sasaki 2000
- Rodina Alariaceae Setchell & Gardner 1925
- Rodina Aureophycaceae Kawai & Ridgway 2013
- Rodina Chordaceae Dumortier 1822
- Rodina Laminariaceae Bory de Saint-Vincent 1827 [Arthrothamnaceae Petrov 1974]
- Rodina Druh Lessoniaceae Setchell & Gardner 1925
- Rodina Pseudochordaceae Kawai a Kurogi 1985
- Objednat Nemodermatales Parente a kol. 2008
- Rodina Nemodermataceae Kuckuck ex Feldmann 1937
- Objednat Phaeosiphoniellales Silberfeld, Rousseau & Reviers 2014
- Rodina Phaeosiphoniellaceae Phillips et al. 2008
- Objednat Ralfsiales Nakamura ex Lim & Kawai 2007
- Rodina Mesosporaceae Tanaka & Chihara 1982
- Rodina Neoralfsiaceae Lim & Kawai 2007
- Rodina Ralfsiaceae Farlow 1881 [Heterochordariaceae Setchell & Gardner 1925]
- Objednat Scytothamnales Vychází Peters & Clayton 1998. Silberfeld a kol. 2011
- Rodina Asteronemataceae Silberfeld a kol. 2011
- Rodina Bachelotiaceae Silberfeld a kol. 2011
- Rodina Splachnidiaceae Mitchell & Whitting 1892 [Scytothamnaceae Womersley 1987]
- Objednat Sporochnales Sauvageau 1926
- Rodina Sporochnaceae Greville 1830
- Objednat Tilopteridales Bessey 1907 vychází. Phillips et al. 2008 [Cutleriales Bessey 1907]
- Rodina Cutleriaceae Griffith & Henfrey 1856
- Rodina Halosiphonaceae Kawai & Sasaki 2000
- Rodina Phyllariaceae Tilden 1935
- Rodina Stschapoviaceae Kawai 2004
- Rodina Tilopteridaceae Kjellman 1890
- Objednat Ascoseirales Petrov 1964 vychází. Moe a Henry 1982
- Podtřída Discosporangiophycidae Silberfeld, Rousseau & Reviers 2014
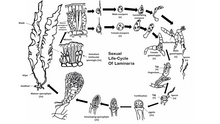
Životní cyklus
Většina hnědých řas, s výjimkou Fucales, provést sexuální reprodukce přes sporová meióza.[48] Mezi generacemi procházejí řasy odděleně sporofyt (diploidní ) a gametofyt (haploidní ) fáze. Fáze sporofytů je často viditelnější z těchto dvou, ačkoli některé druhy hnědých řas mají podobné diploidní a haploidní fáze. Volně plovoucí formy hnědých řas často nepodstupují sexuální reprodukce dokud se nepřichytí k podkladu. Haploidní generace se skládá z muže a ženy gametofyty.[49] Oplodnění vaječných buněk se u různých druhů hnědých řas liší a může být izogamní, oogamní nebo anizogamní. Hnojení může probíhat ve vodě s vejci a pohyblivými spermiemi nebo uvnitř oogonium sám.
Některé druhy hnědých řas mohou také fungovat nepohlavní reprodukce prostřednictvím výroby pohyblivého diploidu zoospory. Tyto zoospory se tvoří v plurilokulárním sporangium a může okamžitě dospět do fáze sporofytů.
U reprezentativního druhu Laminaria, je nápadný diploidní generace a menší haploidní generace. Meióza probíhá během několika unilocular sporangium podél čepele řasy, z nichž každá tvoří buď haploidního muže nebo ženu zoospory. Spory se poté uvolňují ze sporangií a rostou za vzniku mužských a ženských gametofytů. Samice gametofytů produkuje vajíčko v oogoniu a samci gametofyt uvolňují pohyblivé spermie, které vajíčko oplodňují. Oplodněná zygota pak roste do dospělého diploidního sporofytu.
V pořadí Fucales, sexuální reprodukce je oogamní a zralý diploid je jedinou formou pro každou generaci. Gamety se tvoří ve specializovaných conceptacles které se vyskytují rozptýleny na obou površích nádoby, vnější části čepelí mateřské rostliny. Vaječné buňky a pohyblivé spermie se uvolňují ze samostatných vaků v objektech mateřských řas a ve vodě se dokončí oplodnění. Oplodněná zygota se usazuje na povrchu a poté se diferencuje na listovou thallus a prstem připomínající pevnost. Světlo reguluje diferenciaci zygoty na čepel a pevnou polohu.

Ekologie
Hnědé řasy se přizpůsobily široké škále mořských ekologických výklenků, včetně přílivové zóny, skalních bazénů, celé přílivové zóny a relativně hluboko u břehů. Jsou důležitou složkou některých brakických vodních ekosystémů a sladkovodní kolonizovaly maximálně šestkrát.[50] Velké množství Phaeophyceae je přílivové nebo horní litorální,[26] a jsou to převážně studené a studené vodní organismy, které těží z živin v proudu studené vody a přítoků ze země; Sargassum být prominentní výjimkou z tohoto zevšeobecňování.
Hnědé řasy rostoucí v brakických vodách jsou téměř výhradně nepohlavní.[26]
Chemie
| Skupina řas | δ13Rozsah C.[51] |
|---|---|
| HCO3-použitím červené řasy | −22,5 ‰ až −9,6 ‰ |
| CO2- pomocí červených řas | −34,5 ‰ až −29,9 ‰ |
| Hnědé řasy | −20,8 ‰ až −10,5 ‰ |
| Zelené řasy | −20,3 ‰ až −8,8 ‰ |
Hnědé řasy mají a δ13C hodnota v rozmezí od -30,0 ‰ do -10,5 ‰, na rozdíl od červených řas a zelených. To odráží jejich různé metabolické dráhy.[52]
Mají celulózové stěny s kyselinou alginovou a také obsahují polysacharid fukoidan v amorfních částech jejich buněčných stěn. Několik druhů (z Padina) kalcifikovat s aragonit jehly.[26]
Fotosyntetický systém hnědých řas je vyroben z a P700 komplex obsahující chlorofyl a. Jejich plastidy také obsahují chlorofyl c a karotenoidy (nejrozšířenější z nich fucoxanthin ).[53]
Hnědé řasy produkují specifický druh taninu phlorotannins ve větším množství než červené řasy.
Důležitost a použití
Hnědé řasy zahrnují řadu jedlé mořské řasy. Všechny hnědé řasy obsahují kyselina alginová (alginát) v jejich buněčných stěnách, který je komerčně extrahován a používán jako průmyslové zahušťovadlo v potravinách a pro další použití.[54] Jeden z těchto produktů se používá v lithium-iontové baterie.[55] Kyselina alginová se používá jako stabilní součást baterie anoda. Tento polysacharid je hlavní složkou hnědých řas a nenachází se v suchozemských rostlinách.
Kyselina alginová může být také použita v akvakultura. Například kyselina alginová posiluje imunitní systém pstruha duhového. Mladší ryby pravděpodobně přežijí, když dostanou stravu s kyselinou alginovou.[56]
Hnědé řasy včetně kelp postele také každoročně fixují významnou část zemského oxidu uhličitého pomocí fotosyntézy.[57] A mohou v nich ukládat velké množství oxidu uhličitého, což nám může pomoci v boji proti změně klimatu.[58]Sargachromanol G., výpis z Sargassum siliquastrum Bylo prokázáno, že má protizánětlivé účinky.[59]
Jedlé hnědé řasy
Kelp (Laminariales)
| Fucales
| Ectocarpales
|
Viz také
Reference
- ^ A b Medlin, L. K .; et al. (1997). Fylogenetické vztahy „zlatých řas“ (haptophyty, heterokontové chromofyty) a jejich plastidů (PDF). Systematika a evoluce rostlin. 11. 187–219. doi:10.1007/978-3-7091-6542-3_11. hdl:10013 / epic.12690. ISBN 978-3-211-83035-2.
- ^ A b C Lim, B.-L .; Kawai, H .; Hori, H .; Osawa, S. (1986). "Molekulární vývoj 5S ribozomální RNA z červených a hnědých řas". Japonský žurnál genetiky. 61 (2): 169–176. doi:10,1266 / jjg.61.169.
- ^ Kjellman, F. R. (1891). "Phaeophyceae (Fucoideae)". In Engler, A .; Prantl, K. (eds.). Die natürlichen Pflanzenfamilien. 1. Lipsko: Wilhelm Engelmann. 176–192.
- ^ A b Cock, J. Mark; Peters, Akira F .; Coelho, Susana M. (08.08.2011). "Hnědé řasy". Aktuální biologie. 21 (15): R573 – R575. doi:10.1016 / j.cub.2011.05.006. PMID 21820616.
- ^ Hoek, Christiaan; den Hoeck, Hoeck Van; Mann, David; Jahns, H.M. (1995). Řasy: úvod do fykologie. Cambridge University Press. p. 166. ISBN 9780521316873. OCLC 443576944.
- ^ A b C Connor, J .; Baxter, C. (1989). Řasy lesy. Monterey Bay Aquarium. ISBN 978-1-878244-01-7.
- ^ A b Dittmer, H. J. (1964). Fylogeneze a forma v rostlinné říši. Princeton, NJ: Společnost D. Van Nostrand. str.115–137. ISBN 978-0-88275-167-2.
- ^ Abbott, I. A .; Hollenberg, G. J. (1976). Mořské řasy Kalifornie. Kalifornie: Press Stanford University. ISBN 978-0-8047-0867-8.
- ^ Cribb, A. B. (1953). "Macrocystis pyrifera (L.) Ag. v tasmánských vodách “. Australian Journal of Marine and Freshwater Research. 5 (1): 1–34. doi:10.1071 / MF9540001.
- ^ Jones, W. E. (1962). „Klíč k rodům britských mořských řas“ (PDF). Terénní studie. 1 (4): 1–32.[trvalý mrtvý odkaz ]
- ^ A b C d Bold, H. C .; Alexopoulos, C. J .; Delevoryas, T. (1987). Morfologie rostlin a hub (5. vydání). New York: Vydavatelé Harper & Row. str. 112–131, 174–186. ISBN 978-0-06-040839-8.
- ^ Raven, P. H .; Evert, R.F .; Eichhorn, S.E. (2005). Biologie rostlin (7. vydání). New York: W. H. Freeman and Company. str.316 –321, 347. ISBN 978-0-7167-1007-3.
- ^ Round, F. E. (1981). Ekologie řas. Cambridge: Cambridge University Press. p. 103. ISBN 978-0-521-26906-3.
- ^ Wynne, M. J. (1981). "Biologie mořských řas". V Lobban, C. S .; Wynne, M. J. (eds.). Phaeophyta: Morfologie a klasifikace. Botanické monografie. 17. University of California Press. p. 52. ISBN 978-0-520-04585-9.
- ^ A b Sharma, O. P (1986). Učebnice řas. Tata McGraw-Hill. p. 298. ISBN 978-0-07-451928-8.
- ^ Graham; Wilcox; Graham (2009). Algae, 2. vydání. Pearson. ISBN 9780321603128.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Fritsch, F. E. 1945. Struktura a reprodukce řas. Cambridge University Press, Cambridge.
- ^ Charrier, Bénédicte; Le Bail, Aude; de Reviers, Bruno (srpen 2012). „Plant Proteus: morfologická plasticita hnědých řas a základní vývojové mechanismy“. Trendy ve vědě o rostlinách. 17 (8): 468–477. doi:10.1016 / j.tplantts.2012.03.003. ISSN 1360-1385.
- ^ Charrier, B., H. Rabillé a B. Billoud. 2019. Pohled na expanzi buněčné stěny pod zlatým světlem. Trendy ve vědě o rostlinách 24: 130–141.
- ^ Michel, G., T. Tonon, D. Scornet, J. M. Cock a B. Kloareg. 2010. Metabolismus polysacharidů buněčné stěny hnědé řasy Ectocarpus siliculosus. Pohledy na vývoj polysacharidů extracelulární matrix u eukaryot. Nový fytolog 188: 82–97.
- ^ Deniaud-Bouët, E., N. Kervarec, G. Michel, T. Tonon, B. Kloareg a C. Hervé. 2014. Chemická a enzymatická frakcionace buněčných stěn z Fucales: pohledy do struktury extracelulární matrice hnědých řas. Annals of Botany 114: 1203–1216.
- ^ Adl, S. M .; et al. (2005). „Nová vyšší klasifikace eukaryot s důrazem na taxonomii protistů“ (PDF). Journal of Eukaryotic Microbiology. 52 (5): 399–451. doi:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873.
- ^ Lane, C.E .; Archibald, J. M. (2008). „Eukaryotický strom života: Endosymbióza má svůj TOL“ (PDF). Trendy v ekologii a evoluci. 23 (5): 268–275. CiteSeerX 10.1.1.555.2930. doi:10.1016 / j.tree.2008.02.004. PMID 18378040.
- ^ A b van den Hoek, C .; Mann, D. G .; Jahns, H. M. (1995). Algae: An Introduction to Phycology. Cambridge: Cambridge University Press. 165–218. ISBN 978-0-521-31687-3.
- ^ Niklas, K. J. (1997). Evoluční biologie rostlin. Chicago: University of Chicago Press. p. 156. ISBN 978-0-226-58082-1.
- ^ A b C d E Lee, R. E. (2008). Fykologie (4. vydání). Cambridge University Press. ISBN 978-0-521-63883-8.
- ^ Ariztia, E. V .; Andersen, R. A .; Sogin, M. L. (1991). „Nová fylogeneze chromofytových řas využívající rSNA sekvence podobné 16S z Mallomonas papillosa (Synurophyceae) a Tribonema aequale (Xanthophyceae) ". Journal of Phycology. 27 (3): 428–436. doi:10.1111 / j.0022-3646.1991.00428.x.
- ^ Taylor, T. N .; Taylor, E.L. (1993). Biologie a evoluce fosilních rostlin. Englewood Cliffs, NJ: Prentice Hall. str. 128–131. ISBN 978-0-13-651589-0.
- ^ Dittmer, H. J. (1964). Fylogeneze a forma v rostlinné říši. Princeton, NJ: D. Van Nostrand Company. str.115–137. ISBN 978-0-88275-167-2.
- ^ Hori, H .; Osawa, S. (1987). "Původ a vývoj organismů odvozený ze sekvencí 5S ribozomálních RNS" (PDF). Molekulární biologie a evoluce. 4 (5): 445–472. doi:10.1093 / oxfordjournals.molbev.a040455. PMID 2452957.
- ^ A b Arnold, C. A. (1947). Úvod do paleobotaniky. New York; Londýn: McGraw-Hill. p.48. ISBN 978-1-4067-1861-4.
- ^ Coyer, J. A .; Smith, G. J .; Andersen, R. A. (2001). "Vývoj Makrocystis spp. (Phaeophyta), jak je stanoveno sekvencemi ITS1 a ITS2 " (PDF). Journal of Phycology. 37 (4): 574–585. doi:10.1046 / j.1529-8817.2001.037001574.x.
- ^ A b C Fry, W. L .; Banks, H. P. (1955). "Tři nové rody z horního devonu v New Yorku". Journal of Paleontology. 29 (1): 37–44. JSTOR 1300127.
- ^ Prescott, G. W. (1968). The Algae: A Review. Boston: Společnost Houghton Mifflin. str.207 –231, 371–372. ISBN 978-3-87429-244-3.
- ^ Simpson, G. G. (1953). Život minulosti: Úvod do paleontologie. Nové nebe: Yale University Press. str.158–159.
- ^ Fry, W. L. (1983). „Alga flora from the Upper Ordovician of the Lake Winnipeg region, Manitoba, Canada“. Recenze paleobotaniky a palynologie. 39 (3–4): 313–341. doi:10.1016/0034-6667(83)90018-0.
- ^ Speer, B. R .; Wagoner, B. M. (2000). „Phaeophyta: Fosilní záznam“.
- ^ Loeblich, A. R. (1974). "Fylogeneze Protistanu podle fosilního záznamu". Taxon. 23 (2/3): 277–290. doi:10.2307/1218707. JSTOR 1218707.
- ^ Hofmann, H. J. (1985). „Precambrian Carbonaceous Megafossils“. V D. F. Toomey; M. H. Nitecki (eds.). Paleoalgologie: Současný výzkum a aplikace. Berlín: Springer-Verlag. 20–33.
- ^ Hofmann, H. J. (1994). "Proterozoické uhlíkaté komprese (" metafyty "a" červy ")". V Bengtson, S. (ed.). Život na Zemi. Nobelovo sympozium. 84. New York: Columbia University Press. str. 342–357.
- ^ Hueber, F. M. (2001). „Shnilé dřevo-řasy: historie a život Prototaxity Dawson 1859 ". Recenze paleobotaniky a palynologie. 116 (1): 123–158. doi:10.1016 / S0034-6667 (01) 00058-6.
- ^ Taylor, W. A .; Taylor, T. N. (1987). "Sporová stěna ultrastruktury Protosalvinie" (PDF). American Journal of Botany. 74 (3): 437–443. doi:10.2307/2443819. JSTOR 2443819. Archivovány od originál (PDF) dne 2010-06-17.
- ^ Leary, R.L. (1986). "Tři nové rody fosilních nevápnitých řas z Valmeyeran (Mississippian) vrstev Illinois". American Journal of Botany. 73 (3): 369–375. doi:10.2307/2444080. JSTOR 2444080.
- ^ Bold, H. C .; Wynne, M. J. (1978). Úvod do řas (2. vyd.). Prentice-Hall. p.27. ISBN 978-0-13-477786-3.
- ^ Parker, B. C .; Dawson, E. Y. (1965). „Nevápenaté mořské řasy z ložisek kalifornského miocénu“. Nova Hedwigia. 10: 273–295, desky 76–96.
- ^ A b Silberfeld, Thomas; Rousseau, Florencie; de Reviers, Bruno (2014). „Aktualizovaná klasifikace hnědých řas (Ochrophyta, Phaeophyceae)“. Cryptogamie, Algologie. 35 (2): 117–156. doi:10.7872 / crya.v35.iss2.2014.117.
- ^ Guiry, M. D .; Guiry, G. M. (2009). „AlgaeBase“. Irská národní univerzita. Citováno 2012-12-31.
- ^ Bold, Harold Charles; Wynne, Michael James (01.01.1985). Úvod do řas: struktura a reprodukce. Prentice-Hall.
- ^ Lesley., Lovett-Doust, Jon and (1990-01-01). Ekologie reprodukce rostlin: vzory a strategie. Oxford University Press. ISBN 9780198021926. OCLC 437173314.
- ^ Dittami, SM; Heesch, S; Olsen, JL; Collén, J (2017). „Přechody mezi mořským a sladkovodním prostředím poskytují nové informace o původu mnohobuněčných rostlin a řas“ (PDF). J. Phycol. 53 (4): 731–745. doi:10.1111 / jpy.12547. PMID 28509401.
- ^ Maberly, S. C .; Raven, J. A .; Johnston, A. M. (1992). „Diskriminace mezi 12C a 13C mořskými rostlinami ". Ekologie. 91 (4): 481. doi:10.1007 / BF00650320. JSTOR 4220100.
- ^ Fletcher, B. J .; Beerling, D. J.; Chaloner, W. G. (2004). „Stabilní uhlíkové izotopy a metabolismus suchozemského devonského organismu Spongiophyton“. Geobiologie. 2 (2): 107–119. doi:10.1111 / j.1472-4677.2004.00026.x.
- ^ Berkaloff, Claire (1990). „Organizace podjednotek částic PSI z hnědých řas a rozsivek: analýza polypeptidů a pigmentů“. Fotosyntetický výzkum. 23 (2): 181–193. doi:10.1007 / BF00035009. PMID 24421060.
- ^ "Kyselina alginová". www.fao.org. Citováno 2017-04-20.
- ^ Kovalenko, Igor; Zdyrko, Bogdan; Magasinski, Alexandre; Hertzberg, Benjamin; Milicev, Zoran; Burtovyy, Ruslan; Lužinov, Igor; Yushin, Gleb (01.01.2011). „Hlavní složka hnědých řas pro použití ve vysokokapacitních lithium-iontových bateriích“. Věda. 334 (6052): 75–79. Bibcode:2011Sci ... 334 ... 75K. doi:10.1126 / science.1209150. JSTOR 23059304. PMID 21903777.
- ^ Gioacchini, Giorgia; Lombardo, Francesco; Avella, Matteo Alessandro; Olivotto, Ike; Carnevali, Oliana (01.04.2010). "Zlepšení dobrých životních podmínek pomocí kyseliny alginové u mladistvých pstruhů duhových (Oncorhynchus mykiss)". Chemie a ekologie. 26 (2): 111–121. doi:10.1080/02757541003627738. ISSN 0275-7540.
- ^ Vásquez, Julio A .; Zuñiga, Sergio; Tala, Fadia; Piaget, Nicole; Rodríguez, Deni C .; Vega, J. M. Alonso (2014-04-01). „Ekonomické hodnocení řasových lesů v severním Chile: hodnoty zboží a služeb ekosystému“. Journal of Applied Phycology. 26 (2): 1081–1088. doi:10.1007 / s10811-013-0173-6. ISSN 0921-8971.
- ^ Krause-Jensen, D .; Duarte, C. (21. února 2020). "Podstatná role makroras v mořském sekvestraci uhlíku". Nature Geoscience. 9 (10): 737–742. doi:10.1038 / ngeo2790.
- ^ Yoon, Weon-Jong; Heo, Soo-Jin; Han, Sang-Chul; Lee, Hye-Ja; Kang, Gyeoung-Jin; Kang, Hee-Kyoung; Hyun, Jin-Won; Koh, Young-Sang; Yoo, Eun-Sook (01.08.2012). „Protizánětlivý účinek sargachromanolu G izolovaného ze Sargassum siliquastrum v buňkách RAW 264,7“. Archivy farmaceutického výzkumu. 35 (8): 1421–1430. doi:10.1007 / s12272-012-0812-5. ISSN 0253-6269. PMID 22941485.