Aspergillus fumigatus - Aspergillus fumigatus
| Aspergillus fumigatus | |
|---|---|
 | |
| Vědecká klasifikace | |
| Království: | Houby |
| Divize: | Ascomycota |
| Třída: | Eurotiomycetes |
| Objednat: | Eurotiales |
| Rodina: | Trichocomaceae |
| Rod: | Aspergillus |
| Druh: | A. fumigatus |
| Binomické jméno | |
| Aspergillus fumigatus Fresenius 1863 | |
| Synonyma | |
Neosartorya fumigata | |
Aspergillus fumigatus je druh houba v rodu Aspergillus, a je jedním z nejběžnějších Aspergillus druhy způsobující nemoci u jedinců s imunodeficience.
Aspergillus fumigatus, a saprotrof v přírodě rozšířený, obvykle se vyskytuje v půdě a rozpadajících se organických látkách, jako jsou hromady kompostu, kde hraje zásadní roli uhlík a dusík recyklace.[1] Kolonie houby produkují z konidiofory; tisíce minut šedozelené konidie (2–3 μm), které se snadno stanou ve vzduchu. Po mnoho let, A. fumigatus Předpokládalo se, že se rozmnožuje pouze nepohlavně, protože ani páření ani redukční dělení buněk kdy byly pozorovány. V roce 2008, A. fumigatus Bylo prokázáno, že má plně funkční sexuální reprodukční cyklus, 145 let po původním popisu Freseniem.[2] Ačkoli A. fumigatus vyskytuje se v oblastech s velmi odlišným podnebím a prostředím, vykazuje nízkou genetickou variabilitu a nedostatek populační genetické diferenciace v globálním měřítku.[3] Schopnost sexu je tedy zachována, i když se vytváří malá genetická variace.
Houba je schopná růstu při 37 ° C nebo 99 ° F (normální teplota lidského těla ), a může růst při teplotách až 50 ° C nebo 122 ° F, přičemž konidie přežívají při 70 ° C nebo 158 ° F - za podmínek, s nimiž se pravidelně setkává v samohřívacích hromadách kompostu. Jeho výtrusy jsou v atmosféře všudypřítomné a každý každý den vdechuje odhadem několik stovek výtrusů; obvykle jsou tyto u zdravých jedinců rychle eliminovány imunitním systémem. v imunokompromitovaný jednotlivci, jako jsou příjemci transplantovaných orgánů a lidé s AIDS nebo leukémie, houba je pravděpodobnější patogenní, přeběhnutí oslabené obrany hostitele a způsobení řady nemocí obecně nazývaných aspergilóza. Vzhledem k nedávnému nárůstu používání imunosupresiv k léčbě lidských onemocnění se odhaduje, že A. fumigatus může být odpovědný za více než 600 000 úmrtí ročně s úmrtností mezi 25–90%.[4] Několik faktory virulence byly postulovány, aby to vysvětlily oportunistický chování.[5]
Když fermentační vývar z A. fumigatus byl promítán, řada indolický alkaloidy s antimitotický vlastnosti byly objeveny.[6] Sledované sloučeniny patří do třídy známé jako tryprostatiny spirotryprostatin B jako protinádorový lék.
Aspergillus fumigatus pěstované na určitých stavebních materiálech mohou produkovat genotoxický a cytotoxický mykotoxiny, jako gliotoxin.[7]
Genom
Aspergillus fumigatus má stáj haploidní genom 29,4 milionu základní páry. Sekvence genomu tří Aspergillus druh-Aspergillus fumigatus, Aspergillus nidulans, a Aspergillus oryzae —Byly publikovány v Příroda v prosinci 2005.[8][9][10]
Patogeneze
Aspergillus fumigatus je nejčastější příčinou invazivní plísňové infekce u imunosuprimovaných jedinců, mezi něž patří pacienti podstupující imunosupresivní léčbu autoimunitního nebo neoplastického onemocnění, příjemci transplantovaných orgánů a pacienti s AIDS.[11] A. fumigatus primárně způsobuje invazivní infekci v plicích a představuje hlavní příčinu morbidity a mortality u těchto jedinců.[12] Dodatečně, A. fumigatus může způsobit chronické plicní infekce, alergická bronchopulmonální aspergilóza nebo alergické onemocnění u imunokompetentních hostitelů.[13]
Vrozená imunitní odpověď
Inhalační expozice konidiím ve vzduchu je kontinuální kvůli jejich všudypřítomné distribuci v životním prostředí. U zdravých jedinců je však vrozený imunitní systém účinnou překážkou A. fumigatus infekce.[13] Velká část inhalovaných konidií je odstraněna mukociliárním působením respiračního epitelu.[13] Kvůli malé velikosti konidií se mnoho z nich ukládá v alveoly, kde interagují s epiteliálními a vrozenými efektorovými buňkami.[11][13] Alveolární makrofágy fagocytují a ničí konidie ve svých fagosomech.[11][13] Epiteliální buňky, konkrétně pneumocyty typu II, také internalizují konidie, které se přenášejí do lysosomu, kde jsou zničeny požité konidie.[11][13][14] Imunitní buňky první linie také slouží k náboru neutrofily a další zánětlivé buňky uvolňováním cytokiny a chemokiny indukované ligací specifických houbových motivů na receptory pro rozpoznávání patogenů.[13] Neutrofily jsou nezbytné pro rezistenci na aspergilózu, jak je prokázáno u neutropenických jedinců, a jsou schopné sekvestrovat jak konidie, tak hyfy prostřednictvím odlišných nefagocytických mechanismů.[11][12][13] Hyfy jsou příliš velké na internalizaci zprostředkovanou buňkami, a tak poškození vyvolané NADPH-oxidázou zprostředkované neutrofily představuje dominantní obranu hostitele proti hyfám.[11][13] Kromě těchto buněčně zprostředkovaných mechanismů eliminace přispívají k obraně hostitele antimikrobiální peptidy vylučované epitelem dýchacích cest.[11]
Invaze

Imunosuprimovaní jedinci jsou náchylní k invazivitě A. fumigatus infekce, která se nejčastěji projevuje jako invazivní plicní aspergilóza. Inhalační konidie, které se vyhýbají imunitní destrukci hostitele, jsou předky invazivního onemocnění. Tyto konidie vycházejí z klidu a morfologickým přechodem na hyfy klíčí v teplém, vlhkém prostředí plicních alveol bohatých na živiny.[11] Klíčení nastává jak extracelulárně, tak i uvnitř pneumocyty typu II endosomy obsahující konidie.[11][14] Po klíčení má růst vláknitých hyfů za následek penetraci do epitelu a následnou penetraci do vaskulárního endotelu.[11][14] Proces angioinvaze způsobuje poškození endotelu a vyvolává prozánětlivou odpověď, expresi tkáňového faktoru a aktivaci koagulace kaskáda.[11] To má za následek intravaskulární trombóza a lokalizovaná tkáň infarkt, nicméně, šíření hyphal fragmentů je obvykle omezené.[11][14] K šíření prostřednictvím krevního oběhu dochází pouze u jedinců se sníženou imunitou.[14]
Odpověď na hypoxii
Jak je u nádorových buněk a jiných patogenů běžné, invazivní hyfy A. fumigatus se v místě infekce v hostitelském organismu setkávají s hypoxickým (nízké hladiny kyslíku, ≤ 1%) mikroprostředím.[15][16][17] Současný výzkum naznačuje, že po infekci nekróza a zánět způsobují poškození tkání, což snižuje dostupné koncentrace kyslíku v důsledku lokálního snížení perfúze, průchodu tekutin do orgánů. Konkrétně u A. fumigatus bylo zjištěno, že sekundární metabolity inhibují vývoj nových krevních cév, což vede k poškození tkáně, inhibici opravy tkáně a nakonec lokalizovaným hypoxickým mikroprostředím.[16] Přesné důsledky hypoxie na houbovou patogenezi nejsou v současné době známy, avšak toto prostředí s nízkým obsahem kyslíku je již dlouho spojováno s negativními klinickými výsledky. Vzhledem k významným zjištěným korelacím mezi hypoxií, plísňovými infekcemi a negativními klinickými výsledky jsou mechanismy, kterými se A. fumigatus adaptuje na hypoxii, rostoucí oblastí zaměření na nové cíle léků.
Ukázalo se, že dva vysoce charakterizované proteiny vázající sterol-regulační prvek, SrbA a SrbB, spolu s jejich procesními cestami ovlivňují vhodnost A. fumigatus v hypoxických podmínkách. Transkripční faktor SrbA je hlavním regulátorem při houbové reakci na hypoxii in vivo a je nezbytný v mnoha biologických procesech, včetně homeostázy železa, rezistence na fungicidní azol a virulence.[18] V důsledku toho ztráta SrbA vede k neschopnosti A. fumigatus růst za podmínek s nízkým obsahem železa, vyšší citlivosti na antifungální azolové léky a úplné ztrátě virulence u myších modelů IPA (invazivní plicní aspergilóza).[19] SrbA knockout mutanti nevykazují žádné známky in vitro růstu v nízkém obsahu kyslíku, o kterém se předpokládá, že je spojen s oslabenou virulencí. SrbA funkčnost při hypoxii závisí na procesu předřazeného štěpení prováděném proteiny RbdB, SppA a Dsc A-E.[20][21][22] SrbA je štěpen z endoplazmatického retikula, které obsahuje 1015 aminokyselinových prekurzorových proteinů, na funkční formu 381 aminokyselin. Ztráta kteréhokoli z výše uvedených zpracovatelských proteinů SrbA má za následek nefunkční kopii SrbA a následnou ztrátu in vitro růstu hypoxie a oslabenou virulenci. Studie imunoprecipitace chromatinu s proteinem SrbA vedly k identifikaci druhého regulátoru hypoxie, SrbB.[19] Ačkoli je o zpracování SrbB známo jen málo, tento transkripční faktor se také ukázal být klíčovým hráčem virulence a fungální hypoxické odpovědi.[19] Podobně jako SrbA, SrbB knockout mutant vedl ke ztrátě virulence, nicméně nedošlo k žádné zvýšené citlivosti vůči antifungálním lékům ani k úplné ztrátě růstu za hypoxických podmínek (50% snížení SrbB spíše než 100% snížení SrbA).[19][18] Stručně řečeno, SrbA i SrbB se ukázaly jako kritické při adaptaci A. fumigatus u savčího hostitele.
Získávání živin
Aspergillus fumigatus musí získat živiny ze svého vnějšího prostředí, aby přežil a vzkvétal uvnitř svého hostitele. Ukázalo se, že mnoho genů zapojených do těchto procesů ovlivňuje virulenci prostřednictvím experimentů zahrnujících genetické mutace. Mezi příklady příjmu živin patří příjem kovů, dusíku a makromolekul, jako jsou peptidy.[12][23]
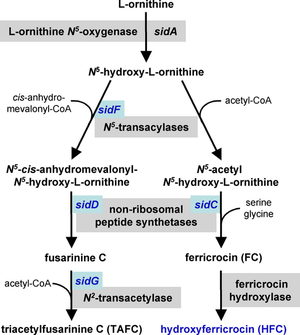
Akvizice železa
Železo je nutné kofaktor pro mnoho enzymů a může působit jako katalyzátor v elektronovém transportním systému. A. fumigatus má dva mechanismy pro příjem železa, redukční získávání železa a siderofor -zprostředkovaný.[25][26] Reduktivní získávání železa zahrnuje přeměnu železa z železitý (Fe+3) do železný (Fe+2) stav a následný příjem přes FtrA, žehličku permeáza. Cílená mutace genu ftrA neindukovala snížení virulence v myší model A. fumigatus invaze. Naproti tomu cílená mutace sidA, prvního genu v dráze biosyntézy sideroforu, se ukázala jako nezbytná pro virulenci sideroforem zprostředkovaného vychytávání železa.[26][27] Mutace následných genů biosyntézy sideroforů sidC, sidD, sidF a sidG vedly k kmenům A. fumigatus s podobným poklesem virulence.[24] Zdá se, že tyto mechanismy vychytávání železa fungují paralelně a oba jsou regulovány v reakci na hladovění železa.[26]
Asimilace dusíku
Aspergillus fumigatus může přežít na různých dusík zdroje a asimilace dusíku má klinický význam, protože bylo prokázáno, že ovlivňuje virulenci.[23][28] Proteiny podílející se na asimilaci dusíku jsou transkripčně regulovány genem AfareA v A. fumigatus. Cílená mutace genu afareA ukázala pokles nástupu úmrtnosti u myšího modelu invaze.[28] The Ras regulovaný protein RhbA se také podílí na asimilaci dusíku. Bylo zjištěno, že RhbA je transkripčně upregulován po kontaktu s A. fumigatus s člověkem endoteliální buňky a kmeny s cílenou mutací rhbA Gen vykazoval snížený růst na špatných zdrojích dusíku a sníženou virulenci in vivo.[29]
Proteinázy
Lidská plíce obsahuje velké množství kolagen a elastin, proteiny, které umožňují flexibilitu tkání.[30] Aspergillus fumigatus produkuje a vylučuje elastázy, proteázy které štěpí elastin, aby rozložily tyto makromolekulární polymery pro absorpci. Významná korelace mezi množstvím produkce elastázy a invazí do tkání byla poprvé objevena v roce 1984.[31] Bylo také zjištěno, že klinické izoláty mají větší aktivitu elastázy než kmeny prostředí A. fumigatus.[32] Byla charakterizována řada elastáz, včetně těch z serinová proteáza, aspartátová proteáza, a metaloproteáza rodiny.[33][34][35][36] Velká redundance těchto elastáz přesto bránila identifikaci specifických účinků na virulenci.[12][23]
Rozložená proteinová odpověď
Řada studií zjistila, že rozvinutá proteinová odpověď přispívá k virulenci A. fumigatus.[37]
Sekundární metabolismus
Sekundární metabolity ve vývoji hub
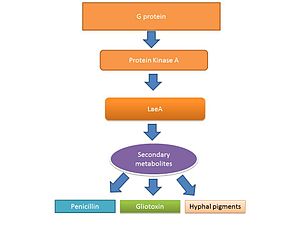
Životní cyklus vláknitých hub včetně Aspergillus spp. sestává ze dvou fází: a hyphal růstová fáze a reprodukční (sporulace ) fáze. Přepínání mezi růstovou a reprodukční fází těchto hub je částečně regulováno úrovní produkce sekundárního metabolitu.[39][40] The sekundární metabolity Předpokládá se, že jsou produkovány k aktivaci sporulace a pigmentů potřebných pro sporulační struktury.[41][42] Signalizace G proteinu reguluje produkci sekundárního metabolitu.[43] Sekvenování genomu odhalilo 40 potenciálních genů zapojených do produkce sekundárních metabolitů, včetně mykotoxinů, které se produkují v době sporulace.[44][45]
Gliotoxin
Gliotoxin je mykotoxin schopný měnit obranyschopnost hostitele prostřednictvím imunosuprese. Neutrofily jsou hlavním cílem gliotoxinu.[46][47] Gliotoxin přerušuje funkci leukocytů inhibicí migrace a produkce superoxidu a způsobuje apoptózu v makrofágech.[48] Gliotoxin narušuje prozánětlivou reakci inhibicí NF-kB.[49]
Transkripční regulace gliotoxinu
LaeA a GliZ jsou transkripční faktory, o nichž je známo, že regulují produkci gliotoxinu. LaeA je univerzální regulátor produkce sekundárního metabolitu v Aspergillus spp.[38] LaeA ovlivňuje expresi 9,5% A. fumigatus genom, včetně mnoha genů biosyntézy sekundárních metabolitů, jako je nonribozomální peptidové syntetázy.[50] Produkce mnoha sekundárních metabolitů, včetně gliotoxinu, byla narušena u kmene LaeA mutantů (AlaEA).[50] Mutant AlaeA vykazoval zvýšenou náchylnost k makrofág fagocytóza a snížená schopnost zabíjet neutrofily ex vivo.[47] Toxiny regulované LaeA kromě gliotoxinu pravděpodobně hrají roli ve virulenci, protože samotná ztráta produkce gliotoxinu rekapituluje hypo-virulentní patotyp ∆laeA.[50]
Současná léčba v boji proti infekcím A. fumigatus
Současná neinvazivní léčba používaná k boji proti plísňovým infekcím sestává z třídy léků známých jako azoly. Azolové léky, jako je vorikonazol, itrakonazol a imidazol, hubí houby tím, že inhibují produkci ergosterolu - kritického prvku buněčných membrán hub. Mechanicky tyto léky působí inhibicí fungálního enzymu cytochromu p450 známého jako 14α-demethyláza.[51] Nicméně, A. fumigatus odolnost vůči azolům se zvyšuje, pravděpodobně v důsledku používání nízkých úrovní azolů v zemědělství.[52][53] Hlavní způsob rezistence je prostřednictvím mutací v cyp51a gen.[54] Byly však pozorovány jiné způsoby rezistence, které představují téměř 40% rezistence u klinických izolátů.[55][56][57] Spolu s azoly existují i jiné třídy fungicidních léků, jako jsou polyeny a echinokandiny. Ačkoli se ukázalo, že tyto léky, zejména azoly, úspěšně bojují proti patogenezi A. fumigatus u imunokompromitovaných a zdravých hostitelů, nové cíle léčiv jsou zásadní kvůli celosvětovému nárůstu kmenů hub rezistentních na léčiva.[58]
Galerie


Conidia phialoconidia of A. fumigatus

Kolonie v Petriho misce

A. fumigatus izolován od lesní půdy

Snímek infikovaného krůtího mozku





Viz také
Reference
- ^ Fang W, Latgé JP (srpen 2018). „Profil mikrobů: Aspergillus fumigatus: saprotrofický a oportunní houbový patogen“. Mikrobiologie. 164 (8): 1009–1011. doi:10,1099 / mic.0.000651. PMC 6152418. PMID 30066670.
- ^ O'Gorman CM, Fuller H, Dyer PS (leden 2009). „Objev sexuálního cyklu v oportunním houbovém patogenu Aspergillus fumigatus“. Příroda. 457 (7228): 471–4. Bibcode:2009Natur.457..471O. doi:10.1038 / nature07528. PMID 19043401.
- ^ Rydholm C, Szakacs G, Lutzoni F (duben 2006). „Nízká genetická variace a žádná zjistitelná struktura populace u aspergillus fumigatus ve srovnání s blízce příbuznými druhy Neosartorya“. Eukaryotická buňka. 5 (4): 650–7. doi:10.1128 / EC. 5.4.450-657.2006. PMC 1459663. PMID 16607012.
- ^ Dhingra S, Cramer RA (2017). „Regulace biosyntézy sterolů v lidském houbovém patogenu Aspergillus fumigatus: Příležitosti pro terapeutický vývoj“. Hranice v mikrobiologii. 8: 92. doi:10.3389 / fmicb.2017.00092. PMC 5285346. PMID 28203225.
- ^ Abad A, Fernández-Molina JV, Bikandi J, Ramírez A, Margareto J, Sendino J a kol. (Prosinec 2010). „Co dělá Aspergillus fumigatus úspěšným patogenem? Geny a molekuly zapojené do invazivní aspergilózy“ (PDF). Revista Iberoamericana de Micologia. 27 (4): 155–82. doi:10.1016 / j.riam.2010.10.003. PMID 20974273.
- ^ Cui CB, Kakeya H, Osada H (srpen 1996). "Spirotryprostatin B, nový inhibitor buněčného cyklu savců produkovaný Aspergillus fumigatus". The Journal of Antibiotics. 49 (8): 832–5. doi:10,7164 / antibiotika. 49,832. PMID 8823522.
- ^ Nieminen SM, Kärki R, Auriola S, Toivola M, Laatsch H, Laatikainen R a kol. (Říjen 2002). „Izolace a identifikace mykotoxinů Aspergillus fumigatus na růstovém médiu a některých stavebních materiálech“. Aplikovaná a environmentální mikrobiologie. 68 (10): 4871–5. doi:10.1128 / aem.68.10.4871-4875.2002. PMC 126391. PMID 12324333.
- ^ Galagan JE, Calvo SE, Cuomo C, Ma LJ, Wortman JR, Batzoglou S a kol. (Prosinec 2005). "Sekvenování Aspergillus nidulans a srovnávací analýza s A. fumigatus a A. oryzae". Příroda. 438 (7071): 1105–15. Bibcode:2005 Natur.438.1105G. doi:10.1038 / příroda04341. PMID 16372000.
- ^ Nierman WC, Pain A, Anderson MJ, Wortman JR, Kim HS, Arroyo J a kol. (Prosinec 2005). "Genomická sekvence patogenní a alergenní vláknité houby Aspergillus fumigatus". Příroda. 438 (7071): 1151–6. Bibcode:2005 Natur.438.1151N. doi:10.1038 / nature04332. PMID 16372009.
- ^ Machida M, Asai K, Sano M, Tanaka T, Kumagai T, Terai G a kol. (Prosinec 2005). "Sekvenování genomu a analýza Aspergillus oryzae". Příroda. 438 (7071): 1157–61. Bibcode:2005 Natur.438.1157M. doi:10.1038 / nature04300. PMID 16372010.
- ^ A b C d E F G h i j k l m Ben-Ami R, Lewis RE, Kontoyiannis DP (Srpen 2010). „Nepřítel stavu (imunosuprese): informace o patogenezi infekce Aspergillus fumigatus“. British Journal of Hematology. 150 (4): 406–17. doi:10.1111 / j.1365-2141.2010.08283.x. PMID 20618330.
- ^ A b C d Hohl TM, Feldmesser M (listopad 2007). "Aspergillus fumigatus: principy patogeneze a obrany hostitele". Eukaryotická buňka. 6 (11): 1953–63. doi:10.1128 / EC.00274-07. PMC 2168400. PMID 17890370.
- ^ A b C d E F G h i Segal BH (duben 2009). „Aspergilóza“. The New England Journal of Medicine. 360 (18): 1870–84. doi:10.1056 / NEJMra0808853. PMID 19403905.
- ^ A b C d E F Filler SG, Sheppard DC (prosinec 2006). "Plísňová invaze normálně nefagocytických hostitelských buněk". PLoS patogeny. 2 (12): e129. doi:10.1371 / journal.ppat.0020129. PMC 1757199. PMID 17196036.
- ^ Grahl N, Cramer RA (únor 2010). „Regulace adaptace hypoxie: přehlížená vlastnost virulence patogenních hub?“. Lékařská mykologie. 48 (1): 1–15. doi:10.3109/13693780902947342. PMC 2898717. PMID 19462332.
- ^ A b Grahl N, Shepardson KM, Chung D, Cramer RA (květen 2012). „Hypoxie a houbová patogeneze: do vzduchu nebo do vzduchu?“. Eukaryotická buňka. 11 (5): 560–70. doi:10.1128 / EC.00031-12. PMC 3346435. PMID 22447924.
- ^ Wezensky SJ, Cramer RA (duben 2011). „Důsledky hypoxických mikroprostředí během invazivní aspergilózy“. Lékařská mykologie. 49 (Suppl 1): S120–4. doi:10.3109/13693786.2010.495139. PMC 2951492. PMID 20560863.
- ^ A b Willger SD, Puttikamonkul S, Kim KH, Burritt JB, Grahl N, Metzler LJ, Barbuch R, Bard M, Lawrence CB, Cramer RA (listopad 2008). „Protein vázající sterol-regulační prvek je vyžadován pro polaritu buněk, adaptaci hypoxie, rezistenci na azolová léčiva a virulenci u Aspergillus fumigatus“. PLoS patogeny. 4 (11): e1000200. doi:10.1371 / journal.ppat.1000200. PMC 2572145. PMID 18989462.
- ^ A b C d Chung D, Barker BM, Carey CC, Merriman B, Werner ER, Lechner BE, Dhingra S, Cheng C, Xu W, Blosser SJ, Morohashi K, Mazurie A, Mitchell TK, Haas H, Mitchell AP, Cramer RA (listopad 2014 ). „ChIP-seq a transkriptomové analýzy in vivo Asergillus fumigatus SREBP SrbA odhalily nový regulátor reakce a virulence houbové hypoxie“. PLoS patogeny. 10 (11): e1004487. doi:10.1371 / journal.ppat.1004487. PMC 4223079. PMID 25375670.
- ^ Dhingra S, Kowalski CH, Thammahong A, Beattie SR, Bultman KM, Cramer RA (2016). „RbdB, kosočtverečná proteáza kritická pro aktivaci SREBP a virulenci u Aspergillus fumigatus“. mSphere. 1 (2). doi:10,1 128 / mSphere 00035-16. PMC 4863583. PMID 27303716.
- ^ Bat-Ochir C, Kwak JY, Koh SK, Jeon MH, Chung D, Lee YW, Chae SK (květen 2016). „Signální peptidová peptidáza SppA se podílí na štěpení proteinů vázajících regulační prvky na sterol a adaptaci hypoxie u Aspergillus nidulans“. Molekulární mikrobiologie. 100 (4): 635–55. doi:10,1111 / mmi.13341. PMID 26822492.
- ^ Willger SD, Cornish EJ, Chung D, Fleming BA, Lehmann MM, Puttikamonkul S, Cramer RA (prosinec 2012). „Pro adaptaci na hypoxii, reakce triazolových léků a houbovou virulenci u Aspergillus fumigatus jsou vyžadovány ortology Dsc.“. Eukaryotická buňka. 11 (12): 1557–67. doi:10.1128 / EC.00252-12. PMC 3536281. PMID 23104569.
- ^ A b C Dagenais TR, Keller NP (červenec 2009). "Patogeneze Aspergillus fumigatus u invazivní aspergilózy". Recenze klinické mikrobiologie. 22 (3): 447–65. doi:10.1128 / CMR.00055-08. PMC 2708386. PMID 19597008.
- ^ A b Schrettl M, Bignell E, Kragl C, Sabiha Y, Loss O, Eisendle M a kol. (Září 2007). "Zřetelné role pro intra- a extracelulární siderofory během infekce Aspergillus fumigatus". PLoS patogeny. 3 (9): 1195–207. doi:10.1371 / journal.ppat.0030128. PMC 1971116. PMID 17845073.
- ^ Haas H (září 2003). „Molekulární genetika biosyntézy a příjmu houbových sideroforů: role sideroforů při příjmu a skladování železa“. Aplikovaná mikrobiologie a biotechnologie. 62 (4): 316–30. doi:10.1007 / s00253-003-1335-2. PMID 12759789.
- ^ A b C Schrettl M, Bignell E, Kragl C, Joechl C, Rogers T, Arst HN a kol. (Listopad 2004). „Biosyntéza sideroforu, ale ne reduktivní asimilace železa, je nezbytná pro virulenci Aspergillus fumigatus“. The Journal of Experimental Medicine. 200 (9): 1213–9. doi:10.1084 / jem.20041242. PMC 2211866. PMID 15504822.
- ^ Hissen AH, Wan AN, Warwas ML, Pinto LJ, Moore MM (září 2005). „Aspergillus fumigatus siderophore biosyntetický gen sidA, kódující L-ornithin N5-oxygenázu, je nutný pro virulenci“. Infekce a imunita. 73 (9): 5493–503. doi:10.1128 / IAI.73.9.5493-5503.2005. PMC 1231119. PMID 16113265.
- ^ A b Hensel M, Arst HN, Aufauvre-Brown A, Holden DW (červen 1998). „Role Aspergillus fumigatus areA gen při invazivní plicní aspergilóze“. Molekulární a obecná genetika. 258 (5): 553–7. doi:10,1007 / s004380050767. PMID 9669338.
- ^ Panepinto JC, Oliver BG, Amlung TW, Askew DS, Rhodes JC (srpen 2002). „Exprese rheb homologu Aspergillus fumigatus, rhbA, je indukována hladověním dusíkem“. Plísňová genetika a biologie. 36 (3): 207–14. doi:10.1016 / S1087-1845 (02) 00022-1. PMID 12135576.
- ^ Rosenbloom J (prosinec 1984). "Elastin: vztah proteinové a genové struktury k nemoci". Laboratorní vyšetřování; Journal of Technical Methods and Pathology. 51 (6): 605–23. PMID 6150137.
- ^ Kothary MH, Chase T, Macmillan JD (leden 1984). „Korelace produkce elastázy některými kmeny Aspergillus fumigatus se schopností vyvolat plicní invazivní aspergilózu u myší“. Infekce a imunita. 43 (1): 320–5. doi:10.1128 / IAI.43.1.320-325.1984. PMC 263429. PMID 6360904.
- ^ Blanco JL, Hontecillas R, Bouza E, Blanco I, Pelaez T, Muñoz P a kol. (Květen 2002). „Korelace mezi indexem aktivity elastázy a invazivností klinických izolátů Aspergillus fumigatus“. Journal of Clinical Microbiology. 40 (5): 1811–3. doi:10.1128 / JCM.40.5.1811-1813.2002. PMC 130931. PMID 11980964.
- ^ Reichard U, Büttner S, Eiffert H, Staib F, Rüchel R (prosinec 1990). "Purifikace a charakterizace extracelulární serinové proteinázy z Aspergillus fumigatus a její detekce ve tkáni". Journal of Medical Microbiology. 33 (4): 243–51. doi:10.1099/00222615-33-4-243. PMID 2258912.
- ^ Markaryan A, Morozova I, Yu H, Kolattukudy PE (červen 1994). „Čištění a charakterizace elastinolytické metaloproteázy z Aspergillus fumigatus a imunoelektronový mikroskopický důkaz sekrece tohoto enzymu houbou napadající myší plíce“. Infekce a imunita. 62 (6): 2149–57. doi:10.1128 / IAI.62.6.2149-2157.1994. PMC 186491. PMID 8188335.
- ^ Lee JD, Kolattukudy PE (říjen 1995). „Molekulární klonování cDNA a genu pro elastinolytickou aspartátovou proteinázu z Aspergillus fumigatus a důkazy o jejím vylučování houbou během invaze do plic hostitele“. Infekce a imunita. 63 (10): 3796–803. doi:10.1128 / IAI.63.10.3796-3803.1995. PMC 173533. PMID 7558282.
- ^ Iadarola P, Lungarella G, Martorana PA, Viglio S, Guglielminetti M, Korzus E a kol. (1998). „Poranění plic a degradace složek extracelulární matrice serinovou proteinázou Aspergillus fumigatus“. Experimentální výzkum plic. 24 (3): 233–51. doi:10.3109/01902149809041532. PMID 9635248.
- ^ Feng X, Krishnan K, Richie DL, Aimanianda V, Hartl L, Grahl N a kol. (Říjen 2011). „Funkce senzoru stresu ER nezávislé na HacA synergizují s kanonickým UPR, aby ovlivnily vlastnosti virulence u Aspergillus fumigatus“. PLoS patogeny. 7 (10): e1002330. doi:10.1371 / journal.ppat.1002330. PMC 3197630. PMID 22028661.
- ^ A b Bok JW, Keller NP (duben 2004). „LaeA, regulátor sekundárního metabolismu u Aspergillus spp.“. Eukaryotická buňka. 3 (2): 527–35. doi:10.1128 / EC.3.2.527-535.2004. PMC 387652. PMID 15075281.
- ^ Calvo AM, Wilson RA, Bok JW, Keller NP (září 2002). „Vztah mezi sekundárním metabolismem a vývojem hub“. Recenze mikrobiologie a molekulární biologie. 66 (3): 447–59, obsah. doi:10.1128 / MMBR.66.3.447-459.2002. PMC 120793. PMID 12208999.
- ^ Tao L, Yu JH (únor 2011). „AbaA a WetA řídí různá stadia vývoje Aspergillus fumigatus“. Mikrobiologie. 157 (Pt 2): 313–26. doi:10.1099 / mic.0.044271-0. PMID 20966095.
- ^ Adams TH, Yu JH (prosinec 1998). "Řízení souřadnic produkce sekundárního metabolitu a nepohlavní sporulace u Aspergillus nidulans". Současný názor v mikrobiologii. 1 (6): 674–7. doi:10.1016 / S1369-5274 (98) 80114-8. PMID 10066549.
- ^ Kawamura C, Tsujimoto T, Tsuge T (leden 1999). „Cílené narušení genu pro biosyntézu melaninu ovlivňuje vývoj konidií a UV toleranci v japonském hruškovém patotypu Alternaria alternata“. Molekulární interakce rostlin a mikrobů. 12 (1): 59–63. doi:10.1094 / MPMI.1999.12.1.59. PMID 9885194.
- ^ Brodhagen M, Keller NP (červenec 2006). "Signální cesty spojující produkci mykotoxinů a sporulaci". Molekulární rostlinná patologie. 7 (4): 285–301. doi:10.1111 / j.1364-3703.2006.00338.x. PMID 20507448.
- ^ Nierman WC, Pain A, Anderson MJ, Wortman JR, Kim HS, Arroyo J a kol. (Prosinec 2005). "Genomická sekvence patogenní a alergenní vláknité houby Aspergillus fumigatus". Příroda. 438 (7071): 1151–6. Bibcode:2005 Natur.438.1151N. doi:10.1038 / nature04332. PMID 16372009.
- ^ Trail F, Mahanti N, Linz J (duben 1995). "Molekulární biologie biosyntézy aflatoxinů". Mikrobiologie. 141 (4): 755–65. doi:10.1099/13500872-141-4-755. PMID 7773383.
- ^ Spikes S, Xu R, Nguyen CK, Chamilos G, Kontoyiannis DP, Jacobson RH a kol. (Únor 2008). „Produkce gliotoxinů v Aspergillus fumigatus přispívá k rozdílům virulence specifických pro hostitele“. The Journal of Infectious Diseases. 197 (3): 479–86. doi:10.1086/525044. PMID 18199036.
- ^ A b Bok JW, Chung D, Balajee SA, Marr KA, Andes D, Nielsen KF a kol. (Prosinec 2006). „GliZ, transkripční regulátor biosyntézy gliotoxinu, přispívá k virulenci Aspergillus fumigatus“. Infekce a imunita. 74 (12): 6761–8. doi:10.1128 / IAI.00780-06. PMC 1698057. PMID 17030582.
- ^ Kamei K, Watanabe A (květen 2005). "Aspergillus mycotoxins and their effect on the host". Lékařská mykologie. 43 Suppl 1: S95-9. doi:10.1080/13693780500051547. PMID 16110799.
- ^ Gardiner DM, Waring P, Howlett BJ (duben 2005). „Třída epipolythiodioxopiperazinu (ETP) fungálních toxinů: distribuce, způsob účinku, funkce a biosyntéza“. Mikrobiologie. 151 (Pt 4): 1021–1032. doi:10.1099 / mic.0.27847-0. PMID 15817772.
- ^ A b C Perrin RM, Fedorova ND, Bok JW, Cramer RA, Wortman JR, Kim HS a kol. (Duben 2007). „Transkripční regulace chemické diverzity u Aspergillus fumigatus podle LaeA“. PLoS patogeny. 3 (4): e50. doi:10.1371 / journal.ppat.0030050. PMC 1851976. PMID 17432932.
- ^ Panackal AA, Bennett JE, Williamson PR (září 2014). „Možnosti léčby invazivní aspergilózy“. Současné možnosti léčby infekčních nemocí. 6 (3): 309–325. doi:10.1007 / s40506-014-0016-2. PMC 4200583. PMID 25328449.
- ^ Berger S, El Chazli Y, Babu AF, Coste AT (06.06.2017). „Aspergillus fumigatus: Důsledek antifungálního použití v zemědělství?“. Hranice v mikrobiologii. 8: 1024. doi:10.3389 / fmicb.2017.01024. PMC 5461301. PMID 28638374.
- ^ Bueid A, Howard SJ, Moore CB, Richardson MD, Harrison E, Bowyer P, Denning DW (říjen 2010). „Azolová antifungální rezistence u Aspergillus fumigatus: 2008 a 2009“. The Journal of Antimicrobial Chemotherapy. 65 (10): 2116–8. doi:10.1093 / jac / dkq279. PMID 20729241.
- ^ Snelders E, Karawajczyk A, Schaftenaar G, Verweij PE, Melchers WJ (červen 2010). "Profil rezistence na azolové změny aminokyselin v Aspergillus fumigatus CYP51A na základě modelování proteinové homologie". Antimikrobiální látky a chemoterapie. 54 (6): 2425–30. doi:10.1128 / AAC.01599-09. PMC 2876375. PMID 20385860.
- ^ Rybak JM, Ge W, Wiederhold NP, Parker JE, Kelly SL, Rogers PD, Fortwendel JR (duben 2019). Alspaugh JA (ed.). „hmg1, výzva paradigmatu klinické rezistence na triazol u Aspergillus fumigatus“. mBio. 10 (2): e00437–19, /mbio/10/2/mBio.00437–19.atom. doi:10,1 128 / mBio.00437-19. PMC 6445940. PMID 30940706.
- ^ Camps SM, Dutilh BE, Arendrup MC, Rijs AJ, Snelders E, Huynen MA a kol. (2012-11-30). „Objev mutace HapE, která způsobuje rezistenci na azol u Aspergillus fumigatus prostřednictvím sekvenování celého genomu a sexuálního křížení“. PLOS ONE. 7 (11): e50034. Bibcode:2012PLoSO ... 750034C. doi:10.1371 / journal.pone.0050034. PMC 3511431. PMID 23226235.
- ^ Furukawa T, van Rhijn N, Fraczek M, Gsaller F, Davies E, Carr P a kol. (Leden 2020). „Negativní komplex kofaktoru 2 je klíčovým regulátorem rezistence na léky u Aspergillus fumigatus“. Příroda komunikace. 11 (1): 427. doi:10.1038 / s41467-019-14191-1. PMC 7194077. PMID 31969561.
- ^ Wiederhold NP (2017). „Antifungální rezistence: současné trendy a budoucí strategie boje“. Infekce a odolnost proti drogám. 10: 249–259. doi:10.2147 / IDR.S124918. PMC 5587015. PMID 28919789.
externí odkazy
- Vznik rezistence na azol u Aspergillus fumigatus a šíření jediného mechanismu rezistence. na SciVee
- Důvěra Aspergillus Registrovaná britská charitativní organizace zabývající se celosvětově podporou osob trpících aspergilovou chorobou a výzkumem léčení
- Fungal Research Trust
- Aspergillus informace od DoctorFungus.org