Nervové koreláty vědomí - Neural correlates of consciousness
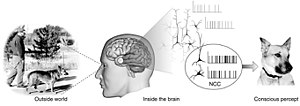
The neurální koreláty vědomí (NCC) tvoří minimální soubor neuronálních událostí a mechanismů dostatečných pro konkrétní vědomí vnímat.[2] Neurologové používají k objevování empirické přístupy neurální koreláty subjektivních jevů; to znamená nervové změny, které nutně a pravidelně korelují s konkrétním zážitkem.[3][4] Sada by měla být minimální protože za předpokladu, že mozek je dostatečný k vyvolání jakékoli dané vědomé zkušenosti, je otázkou, která z jeho složek je nezbytná k jeho výrobě.
Neurobiologický přístup k vědomí
Věda o vědomí musí vysvětlit přesný vztah mezi subjektivními duševními stavy a mozkovými stavy, povahu vztahu mezi vědomou myslí a elektrochemickými interakcemi v těle (problém mysli a těla ). Pokrok v neuropsychologie a neurofilosofie pochází ze soustředění spíše na tělo než na mysl. V této souvislosti lze na neuronální koreláty vědomí pohlížet jako na jejich příčiny a vědomí může být považována za stavově závislou vlastnost nějakého nedefinovaného komplex adaptivní a vysoce propojený biologický systém.[5]
Objevování a charakterizace neurálních korelátů nenabízí teorii vědomí, která by mohla vysvětlit, jak konkrétní systémy vůbec něco zažívají, nebo jak a proč jsou spojeny s vědomím, tzv. těžký problém vědomí,[6] ale pochopení NCC může být krokem k takové teorii. Většina neurobiologů předpokládá, že proměnné, které vedou k vědomí, lze nalézt na neuronové úrovni, ovládané klasickou fyzikou, ačkoli několik vědců navrhlo teorie kvantové vědomí na základě kvantová mechanika.[7]
V neuronových sítích existuje velká zjevná redundance a paralelismus, takže zatímco aktivita v jedné skupině neuronů může v jednom případě korelovat s vnímáním, jiná populace může zprostředkovat související vnímání, pokud dojde ke ztrátě nebo deaktivaci původní populace. Je možné, že každý fenomenální, subjektivní stav má neurální korelaci. Tam, kde lze NCC vyvolat uměle, subjekt zažije přidružené vnímání, zatímco rušení nebo inaktivace oblasti korelace pro konkrétní vnímání ovlivní vnímání nebo způsobí jeho zmizení, což dá vztah příčina-účinek z neurální oblasti k přírodě vnímání.
Co charakterizuje NCC? Jaké jsou společné rysy mezi NCC pro vidění a pro slyšení? Bude NCC zahrnovat všechny pyramidové neurony v kůře v daném okamžiku? Nebo jen podmnožina projekčních buněk s dlouhým dosahem v čelních lalocích, které vyčnívají do senzorických kortik v zadní části? Neurony, které střílejí rytmickým způsobem? Neurony, které střílejí v synchronním způsobem ? Toto jsou některé z návrhů, které byly v průběhu let pokročeny.[8]
Rostoucí schopnost neurologů manipulovat s neurony pomocí metod z molekulární biologie v kombinaci s optickými nástroji (např. Adamantidis et al. 2007 ) závisí na současném vývoji vhodných testů chování a modelových organismů, které lze podrobit genomové analýze a manipulaci ve velkém měřítku. Je to kombinace takové jemnozrnné neuronální analýzy u zvířat se stále citlivějšími psychofyzikálními a mozkovými zobrazovacími technikami u lidí, doplněná vývojem robustního teoretického prediktivního rámce, který snad povede k racionálnímu pochopení vědomí, jednoho z ústřední tajemství života.
Úroveň vzrušení a obsah vědomí
Existují dvě běžné, ale odlišné dimenze termínu vědomí,[9] jeden zahrnující vzrušení a stavy vědomí a další zahrnující obsah vědomí a vědomé stavy. Být při vědomí z cokoli, co mozek musí být v relativně vysokém stavu vzrušení (někdy nazývaného bdělost), ať už v bdělosti nebo REM spánek, živě prožívané ve snech, i když si obvykle nepamatují. Úroveň vzrušení mozku kolísá v a cirkadiánní rytmus, ale může být ovlivněn nedostatkem spánku, drogami a alkoholem, fyzickou námahou atd. Vzrušení lze měřit behaviorálně pomocí amplitudy signálu, která spouští reakci určitého kritéria (například hladina zvuku nutná k vyvolání pohybu očí nebo otočení hlavy) směrem ke zdroji zvuku). Lékaři používají skórovací systémy, jako je Měřítko kómy v Glasgow posoudit úroveň vzrušení u pacientů.
Stavy vysokého vzrušení jsou spojeny s vědomými stavy, které mají konkrétní obsah, vidí, slyší, pamatují, plánují nebo fantazírují o něčem. Různé úrovně nebo stavy vědomí jsou spojeny s různými druhy vědomých zážitků. Stav „vzhůru“ je zcela odlišný od stavu „snění“ (ten druhý má například malou nebo žádnou sebereflexi) a od stavu hlubokého spánku. Ve všech třech případech je ovlivněna základní fyziologie mozku, stejně jako v změněné stavy vědomínapříklad po požití drog nebo během meditace, kdy může dojít ke zlepšení vědomého vnímání a vhledu ve srovnání s běžným stavem bdění.
Lékaři o tom mluví zhoršené stavy vědomí jako v " komatózní stav "," přetrvávající vegetativní stav „(PVS) a„ minimálně vědomý stav „(MCS). Pojem„ stav “zde označuje různá„ množství “vnějšího / fyzického vědomí, od úplné absence v kómatu, přetrvávajícího vegetativního stavu a celkové anestezii, až po kolísavou a omezenou formu vědomí při minimálním vědomí. například při spánkové chůzi nebo během složité částečné epileptický záchvat.[10] Repertoár stavů vědomí nebo zkušeností přístupných pacientovi ve stavu minimálního vědomí je poměrně omezený. Při mozkové smrti nevzniká žádné vzrušení, ale není známo, zda byla přerušena subjektivita zážitku, spíše než jeho pozorovatelné spojení s organismem. Funkční neuroimaging ukázal, že části mozkové kůry jsou stále aktivní u vegetativních pacientů, u nichž se předpokládá, že jsou v bezvědomí;[11] tyto oblasti se však zdají být funkčně odpojeny od asociativních kortikálních oblastí, jejichž aktivita je nutná pro uvědomění.
Potenciál bohatství vědomého zážitku Zdá se, že roste od hlubokého spánku k ospalosti k plné bdělosti, což lze kvantifikovat pomocí představ z teorie složitosti, která zahrnuje jak dimenzionálnost, tak zrnitost vědomého zážitku, aby poskytla integrovaný-informační-teoretický účet vědomí.[12] Jak se vzrušení chování zvyšuje, zvyšuje se také rozsah a složitost možného chování. Přesto v REM spánku existuje charakteristika atonia, nízké motorické vzrušení a člověk se obtížně probouzí, ale stále existuje vysoká metabolická a elektrická mozková aktivita a živé vnímání.
Mnoho jader se zřetelnými chemickými podpisy v thalamus, střední mozek a pons musí fungovat, aby byl subjekt v dostatečném stavu vzrušení mozku, aby mohl vůbec něco zažít. Tato jádra proto patří k umožňujícím faktorům vědomí. Naopak je pravděpodobné, že konkrétní obsah jakéhokoli konkrétního vědomí je zprostředkován určitými neurony v kůře a jejich přidruženými satelitními strukturami, včetně amygdala, thalamus, klaustrum a bazální ganglia.
Neuronální základ vnímání
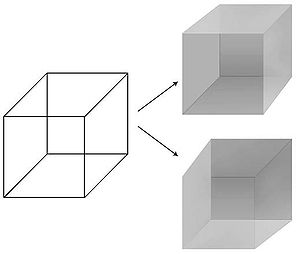
Byla vytvořena možnost přesné manipulace s vizuálními vjemy v čase a prostoru vidění preferovaná modalita při hledání NCC. Psychologové zdokonalili řadu technik - maskování, binokulární soupeření, nepřetržité potlačení blesku, pohybem indukovaná slepota, změnit slepotu, nepozorná slepota - ve kterém je narušen zdánlivě jednoduchý a jednoznačný vztah mezi fyzickým stimulem ve světě a jeho přidruženým vnímáním v soukromí mysli subjektu.[13] Zejména lze stimul vnímavě potlačit po dobu několika sekund nebo dokonce minut: obraz se promítá do jednoho z očí pozorovatele, ale je neviditelný, neviditelný. Tímto způsobem lze izolovat nervové mechanismy, které reagují spíše na subjektivní vnímání než na fyzický stimul, což umožňuje sledování vizuálního vědomí v mozku. V vnímavý iluze, fyzický stimul zůstává pevný, zatímco vnímání kolísá. Nejznámějším příkladem je Neckerova kostka jehož 12 řádků lze do hloubky vnímat jedním ze dvou různých způsobů.
Percepční iluze, kterou lze přesně ovládat, je binokulární soupeření. Zde je levému oku představen malý obraz, např. Vodorovná mřížka, a další obraz, např. Svislá mřížka, je zobrazen na odpovídajícím místě v pravém oku. Navzdory neustálému vizuálnímu podnětu pozorovatelé vědomě vidí, že se horizontální mřížka střídá každých několik sekund s vertikální. Mozek neumožňuje současné vnímání obou obrazů.
Logothetis a kolegové[15] zaznamenal u vzhůru opic makaků provádějících úkol binokulárního soupeření řadu vizuálních kortikálních oblastí. Opice makaků mohou být trénovány, aby hlásily, zda vidí levý nebo pravý obraz. Rozložení spínacích časů a způsob, jakým ovlivňuje změna kontrastu v jednom oku, nezanechává žádné pochybnosti o tom, že opice i lidé mají stejný základní jev. V primární zrakové kůře (V1) pouze malá část buněk slabě modulovala svou odpověď jako funkci vnímání opice, zatímco většina buněk reagovala na jeden nebo druhý podnět sítnice s malým ohledem na to, co zvíře v té době vnímalo. Ale v kortikální oblasti vysoké úrovně, jako je dolní temporální kůra podél ventrální proud téměř všechny neurony reagovaly pouze na percepčně dominantní stimul, takže „obličejová“ buňka vystřelila, pouze když zvíře naznačilo, že vidí obličej a ne vzor prezentovaný druhému oku. To znamená, že NCC zahrnuje neurony aktivní v dolní temporální kůře: je pravděpodobné, že jsou nutné specifické vzájemné akce neuronů v dolní temporální a částech prefrontální kůry.
Počet fMRI experimenty, které využily binokulární rivalitu a související iluze k identifikaci hemodynamické aktivity, která je základem vizuálního vědomí u lidí, zcela přesvědčivě ukazují, že aktivita v horních fázích ventrální dráhy (např. fusiformní oblast obličeje a oblast parahippocampálního místa ) stejně jako v časných oblastech, včetně V1 a laterálního geniculárního jádra (LGN), sledujte percepci, nikoli retinální stimul.[16] Dále řada fMRI[17][18] a DTI experimenty[19] naznačují, že V1 je nezbytná, ale není dostatečná pro vizuální vědomí.[20]
V souvisejícím vnímavém jevu potlačení blesku, vnímání spojené s obrazem promítaným do jednoho oka je potlačeno blikáním jiného obrazu do druhého oka, zatímco původní obraz zůstane. Jeho metodickou výhodou oproti binokulární rivalitě je, že načasování percepčního přechodu je určováno spíše externím spouštěčem než vnitřní událostí. Většina buněk v dolní spánkové kůře a v horním spánkovém žlábku opic trénovaných k hlášení jejich vnímání během bleskové suprese sleduje vnímání zvířete: když je vnímán preferovaný stimul buňky, buňka reaguje. Pokud je obraz stále přítomný na sítnici, ale je percepčně potlačen, buňka ztichne, přestože primární neurony neuronu zrakové kůry vystřelí.[21][22] Záznamy jednotlivých neuronů ve středním temporálním laloku pacientů s epilepsií během bleskové suprese rovněž ukazují zrušení odpovědi, když je preferovaný stimul přítomen, ale percepčně maskovaný.[23]
Globální poruchy vědomí
Vzhledem k absenci jakéhokoli akceptovaného kritéria minimálních neuronových korelátů nezbytných pro vědomí, je rozdíl mezi trvale vegetativním pacientem, který vykazuje pravidelné přechody spánkové vlny a může se pohybovat nebo usmívat, a minimálně vědomým pacientem, který dokáže komunikovat (příležitostně ) smysluplným způsobem (například rozdílnými pohyby očí) a který vykazuje určité známky vědomí, je často obtížné. V globální anestezii by pacient neměl mít psychické trauma, ale úroveň vzrušení by měla být kompatibilní s klinickými potřebami.

Závisí na hladině kyslíku v krvi fMRI prokázali normální vzorce mozkové aktivity u pacienta ve vegetativním stavu po těžkém traumatickém poranění mozku, když byli požádáni, aby si představili hraní tenisu nebo návštěvu pokojů v jeho domě.[25] Diferenciální zobrazování mozku pacientů s takovými globálními poruchami vědomí (včetně akinetický mutismus ) odhalují, že dysfunkce v rozšířené kortikální síti zahrnující mediální a laterální prefrontální a parietální asociační oblasti je spojena s globální ztrátou vědomí.[26] Zhoršené vědomí v epileptický záchvaty temporální lalok bylo rovněž doprovázeno snížením průtoku krve mozkem ve frontální a parietální asociační kůře a zvýšením středních struktur, jako je mediodorsální thalamus.[27]
Relativně lokální bilaterální poranění subkortikálních struktur střední linie (paramediánů) může také způsobit úplnou ztrátu vědomí.[28] Tyto struktury tedy umožnit a řídit vzrušení mozku (jak je určeno metabolickou nebo elektrickou aktivitou) a jsou nezbytnými nervovými koreláty. Jedním z takových příkladů je heterogenní soubor více než dvou desítek jader na každé straně horního mozkového kmene (pons, střední mozek a v zadním hypotalamu), souhrnně označovaný jako retikulární aktivační systém (RAS). Jejich axony se široce promítají do celého mozku. Tato jádra - trojrozměrné sbírky neuronů s vlastní cytoarchitekturou a neurochemickou identitou - uvolňují odlišné neuromodulátory, jako je acetylcholin, noradrenalin / norepinefrin, serotonin, histamin a orexin / hypocretin, které řídí dráždivost thalamu a předního mozku a zprostředkovávají střídání mezi bdělost a spánek i obecná úroveň chování a vzrušení mozku. Po takovém traumatu se však nakonec může vzrušivost thalamu a předního mozku obnovit a vědomí se může vrátit.[29] Dalším faktorem umožňujícím vědomí je pět nebo více intralaminární jádra (ILN) thalamu. Tito přijímají vstup z mnoha jader mozkového kmene a silně se promítají přímo do bazálních ganglií a distribuovanějším způsobem do vrstvy I velké části neokortexu. Poměrně malý (1 cm3 nebo méně) bilaterální léze v talamické ILN úplně vyřadily veškeré vědomí.[30]
Projekce vpřed versus zpětná vazba
Mnoho akcí v reakci na smyslové vstupy je rychlé, přechodné, stereotypní a v bezvědomí.[31] Mohou být považovány za kortikální reflexy a jsou charakterizovány rychlými a poněkud stereotypními odpověďmi, které mohou mít podobu poměrně složitého automatizovaného chování, jak je vidět, např. Ve složitých částečných epileptický záchvaty. Tyto automatizované odpovědi, někdy nazývané zombie chování,[32] může být v kontrastu s pomalejším, víceúčelovým vědomým režimem, který se pomaleji zabývá širšími, méně stereotypními aspekty smyslových vstupů (nebo jejich odrazem, jako v obrazech) a vyžaduje čas, aby se rozhodl o vhodných myšlenkách a odpovědích. Bez takového režimu vědomí by bylo nutné reagovat na neobvyklé události obrovské množství různých režimů zombie.
Vlastností, která odlišuje člověka od většiny zvířat, je to, že se nenarodíme s rozsáhlým repertoárem behaviorálních programů, které by nám umožnily přežít na vlastní pěst („fyziologická předčasnost "). Abychom to vykompenzovali, máme bezkonkurenční schopnost učit se, tj. Vědomě získávat takové programy napodobováním nebo zkoumáním. Jakmile jsou tyto programy vědomě získány a dostatečně procvičeny, mohou se stát automatizovanými do té míry, že jejich provádění probíhá mimo sféry. Vezměte si jako příklad neuvěřitelnou jemnou motoriku při hraní na Beethovenovu klavírní sonátu nebo senzomotorickou koordinaci potřebnou k jízdě na motocyklu po křivolaké horské silnici. Takové složité chování je možné jen proto, že dostatečný počet podprogramů mohou být prováděny s minimálním nebo dokonce pozastaveným vědomým ovládáním. Ve skutečnosti může vědomý systém do jisté míry rušit tyto automatizované programy.[33]
Z evolučního hlediska jasně dává smysl mít jak automatizované behaviorální programy, které lze provádět rychle stereotypně a automatizovaně, tak trochu pomalejší systém, který poskytuje čas na přemýšlení a plánování složitějšího chování. Tento druhý aspekt může být jednou z hlavních funkcí vědomí. Jiní filozofové však navrhli, že vědomí by nebylo nutné pro žádnou funkční výhodu v evolučních procesech.[34][35] Tvrdí, že nikdo nepodal kauzální vysvětlení, proč by pro funkčně ekvivalentní neuvědomělý organismus (tj. filozofický zombie ) dosáhnout stejných výhod přežití jako vědomý organismus. Pokud jsou evoluční procesy slepé k rozdílu mezi funkcemi F provádí vědomý organismus Ó a neuvědomělý organismus Ó*, není jasné, jaké adaptivní výhody by vědomí mohlo poskytnout.[36] Výsledkem je, že exaptivní vysvětlení vědomí získalo přízeň u některých teoretiků, že předpokládané vědomí se nevyvíjelo jako adaptace, ale bylo exaptation vznikající v důsledku dalšího vývoje, jako je zvětšení velikosti mozku nebo kortikální přesmyk.[37] Vědomí v tomto smyslu bylo přirovnáváno k mrtvému bodu v sítnici, kde se nejedná o adaptaci sítnice, nýbrž jen o vedlejší produkt způsobu zapojení sítnicových axonů.[38] Několik vědců včetně Růžovější, Chomsky, Edelman, a Luria naznačili význam vzniku lidského jazyka jako důležitého regulačního mechanismu učení a paměti v kontextu rozvoje vědomí vyššího řádu.
Zdá se možné, že vizuální zombie režimy v kůře používají hlavně hřbetní proud v temenní oblasti.[31] Temenní aktivita však může ovlivnit vědomí tím, že alespoň za určitých okolností vyvolá pozornost na ventrální proud. Vědomý režim vidění závisí do značné míry na časných vizuálních oblastech (mimo V1) a zejména na ventrálním proudu.
Zdánlivě složité vizuální zpracování (například detekce zvířat v přirozených přeplněných scénách) může lidská kůra dosáhnout během 130–150 ms,[39][40] příliš krátké na to, aby došlo k pohybům očí a vědomému vnímání. Kromě toho reflexy, jako je okulovestibulární reflex probíhá v ještě rychlejších časových měřítcích. Je docela pravděpodobné, že takové chování je zprostředkováno čistě dopřednou pohybující se vlnou spiking aktivity, která prochází ze sítnice přes V1, do V4, IT a prefrontální kůry, dokud neovlivňuje motorneurony v míše, které ovládají stisk prstu ( jako v typickém laboratorním experimentu). Hypotéza, že základní zpracování informací je dopředná, je nejpříměji podporována krátkými časy (přibližně 100 ms) potřebnými pro to, aby se v buňkách IT objevila selektivní odpověď.
Naopak se předpokládá, že vědomé vnímání vyžaduje trvalejší dozvukovou nervovou aktivitu, nejpravděpodobněji prostřednictvím globální zpětné vazby z čelních oblastí neokortexu zpět do smyslových kortikálních oblastí[20] který se časem vytváří, dokud nepřekročí kritickou hranici. V tomto okamžiku se vytrvalá nervová aktivita rychle šíří do temenních, prefrontálních a předních cingulárních kortikálních oblastí, thalamu, klaustra a souvisejících struktur, které podporují krátkodobou paměť, multimodální integraci, plánování, řeč a další procesy úzce související s vědomím. Konkurence brání tomu, aby bylo současně a aktivně zastoupeno více než jeden nebo velmi malý počet vjemů. Toto je základní hypotéza teorie globálního pracovního prostoru vědomí.[41][42]
Stručně řečeno, zatímco rychlá, ale přechodná nervová aktivita v thalamokortikálním systému může zprostředkovat komplexní chování bez vědomí, je předpokládáno, že vědomí vyžaduje trvalou, ale dobře organizovanou nervovou aktivitu závislou na kortikokortikální zpětné vazbě na velké vzdálenosti.
Dějiny
Neurobiolog Christfried Jakob (1866-1956) tvrdil, že jediné podmínky, které musí mít neurální koreláty, jsou přímé pocity a reakce; tito se nazývají "intonace".[Citace je zapotřebí ]
Neurofyziologické studie na zvířatech poskytly určité poznatky o nervových korelátech vědomého chování. Vernon Mountcastle počátkem 60. let zahájil studium tohoto souboru problémů, které nazval „problém mysli / mozku“, studiem neurální základny vnímání v somatický senzorický systém. Jeho laboratoře v Johns Hopkins byly mezi prvními, spolu s Edwardem V. Evartsem v NIH, které zaznamenávaly nervovou aktivitu od chovaných opic. V úžasu s elegancí SS Stevensova přístupu odhadu velikosti objevila Mountcastlova skupina tři různé způsoby somatického vjemu, které sdílely jeden kognitivní atribut: ve všech případech rychlost střelby periferních neuronů byla lineárně spojena se silou vyvolaného vnímání. Více nedávno to ukázali Ken H. Britten, William T. Newsome a C. Daniel Salzman oblast MT u opic reagují neurony s variabilitou, která naznačuje, že jsou základem rozhodování o směru pohybu. Nejprve ukázali, že neuronové rychlosti predikují rozhodnutí pomocí teorie detekce signálu, a poté, že stimulace těchto neuronů by mohla předvídatelně ovlivnit rozhodnutí. Po těchto studiích sledoval Ranulfo Romo v somatickém smyslovém systému, aby pomocí odlišného vnímání a oblasti mozku potvrdil, že malý počet neuronů v jedné oblasti mozku je základem percepčních rozhodnutí.
Jiné laboratorní skupiny následovaly Mountcastlovu klíčovou práci týkající se kognitivních proměnných s neuronální aktivitou se složitějšími kognitivními úkoly. Ačkoli opice nemohou mluvit o svém vnímání, byly vytvořeny behaviorální úkoly, při nichž zvířata vytvářela neverbální hlášení, například produkcí pohybů rukou. Mnoho z těchto studií využívá vnímání iluzí jako způsob, jak oddělit pocity (tj., smyslové informace, které mozek přijímá) z vnímání (tj., jak je vědomí interpretuje). Neuronální vzorce, které představují vnímání, nikoli pouze smyslový vstup, jsou interpretovány jako odrážející neuronální korelaci vědomí.
Pomocí takového designu Nikos Logothetis a kolegové objevili neurony odrážející vnímání v temporálním laloku. Vytvořili experimentální situaci, ve které byly konfliktní obrazy prezentovány různým očím (tj., binokulární soupeření ). Za takových podmínek hlásí lidské subjekty bistabilní vjemy: vnímají alternativně jeden nebo druhý obraz. Logothetis a kolegové vycvičili opice, aby hlásily pohyby svých paží, jaký obraz vnímají. Neurony spánkového laloku v experimentech s Logothetis často odrážely to, co opice vnímaly. Neurony s takovými vlastnostmi byly méně často pozorovány v primární zrakové kůře, což odpovídá relativně raným fázím vizuálního zpracování. Další soubor experimentů využívajících binokulární rivalitu u lidí ukázal, že určité vrstvy mozkové kůry lze vyloučit jako kandidáty nervového korelátu vědomí. Logothetis a kolegové si během vnímání jednoho ze snímků vyměnili obrazy mezi očima. Překvapivě zůstalo vnímání stabilní. To znamená, že vědomé vnímání zůstalo stabilní a zároveň se změnil primární vstup do vrstvy 4, což je vstupní vrstva, ve vizuální kůře. Proto vrstva 4 nemůže být součástí nervového korelátu vědomí. Michail Lebedev a jejich kolegové pozorovali podobný jev v opičí prefrontální kůře. Ve svých experimentech opice uváděly vnímaný směr pohybu vizuálních stimulů (což by mohla být iluze) prováděním pohybů očí. Některé neurony prefrontální kůry představovaly skutečné a jiné představovaly vnímané posuny stimulu. Pozorování neuronů souvisejících s vnímáním v prefrontální kůře je v souladu s teorií Christof Koch a Francis Crick kdo předpokládal, že neurální korelát vědomí spočívá v prefrontální kůře. Zastánci distribuovaného zpracování neuronů mohou pravděpodobně zpochybňovat názor, že vědomí má přesnou lokalizaci v mozku.
Francis Crick napsal populární knihu, “Úžasná hypotéza „, jehož tezí je, že neurální korelace vědomí spočívá v našich nervových buňkách a jejich přidružených molekulách. Crick a jeho spolupracovník Christof Koch [43] se snažili vyhnout filozofickým debatám spojeným se studiem vědomí tím, že zdůrazňovali hledání „korelace“ a nikoli „příčinné souvislosti“.[potřebuje aktualizaci ]
O povaze tohoto korelátu existuje velký prostor pro neshody (např., vyžaduje synchronní hroty neuronů v různých oblastech mozku? Je nutná společná aktivace čelních nebo temenních oblastí?). Filozof David Chalmers tvrdí, že neurální korelát vědomí, na rozdíl od jiných korelátů, jako je paměť, nedokáže uspokojivě vysvětlit tento jev; on tomu říká těžký problém vědomí.[44][45]
Viz také
- Vědomí zvířat
- Umělé vědomí
- Mostní lokus
- Poznávací mapa
- Koncepční prostor
- Teorie globálního pracovního prostoru
- Těžký problém vědomí
- Teorie vědomí vyššího řádu
- Schéma obrázku
- Integrovaná teorie informací
- LIDA (kognitivní architektura)
- Modely neurálního výpočtu
- Model s více koncepty
- Münchhausen trilemma
- Neurální kódování
- Neurální dekódování
- Neurální substrát
- Filozofie mysli
- Kvantové poznání
- Kvantová mysl
Poznámky
- ^ Koch 2004, Obrázek 1.1 Neuronální korelace vědomí p. 16.
- ^ Koch 2004, str. 304.
- ^ Vidět tady Archivováno 2013-03-13 na Wayback Machine pro slovník souvisejících pojmů.
- ^ Chalmers, David J. (Červen 1998), „Co je neurální korelát vědomí?“, In Metzinger, Thomas (vyd.), Neurální korelace vědomí: empirické a koncepční otázky, MIT Press (publikováno září 2000), ISBN 978-0-262-13370-8
- ^ Squire 2008, str. 1223.
- ^ Kandel 2007, str. 382.
- ^ Schwartz, Jeffrey M.; Stapp, Henry P.; Beauregard, Mario. „Kvantová fyzika v neurovědě a psychologii: neurofyziologický model interakce mysli a mozku“ (PDF).
- ^ Vidět Chalmers 1998, dostupný online.
- ^ Zeman 2001
- ^ Schiff 2004
- ^ Laureys, Trends Cogn Sci, 2005, 9: 556-559
- ^ Tononi a kol. 2016
- ^ Kim a Blake 2004
- ^ Koch 2004, Obrázek 16.1 Bistabilní Neckerova kostka, str. 270.
- ^ Logothetis 1998
- ^ Rees a Frith 2007
- ^ Haynes a Rees 2005
- ^ Lee a kol. 2007
- ^ Shimono a Niki 2013
- ^ A b Crick a Koch 1995
- ^ Leopold a Logothetis 1996
- ^ Sheinberg a Logothetis 1997
- ^ Kreiman a kol. 2002
- ^ Koch 2004, Obrázek 5.1 Cholinergní povolovací systém p. 92. Viz kapitola 5, která je k dispozici online.
- ^ Owen a kol. 2006
- ^ Laureys 2005
- ^ Blumenfeld a kol. 2004
- ^ Koch 2004, str. 92
- ^ Villablanca 2004
- ^ Bogen 1995
- ^ A b Milner a Goodale 1995
- ^ Koch a Crick 2001
- ^ Beilock a kol. 2002
- ^ Flanagan, Owene; Polger, Tom W. (1995). „Zombie a funkce vědomí“. Journal of Consciousness Studies. 2: 313–321.
- ^ Rosenthal, David (2008). "Vědomí a jeho funkce". Neuropsychologie. 46 (3): 829–840. doi:10.1016 / j.neuropsychologia.2007.11.012. PMID 18164042.
- ^ Harnad, Stevan (2002). „Turingova nerozeznatelnost a nevidomý hodinář“. v Fetzer, James H. (vyd.). Vědomí se vyvíjí. John Benjamins. Citováno 2011-10-26.
- ^ Feinberg, T.E .; Mallatt, J. (2013). „Evoluční a genetické počátky vědomí v kambrijském období před více než 500 miliony let“. Přední Psychol. 4: 667. doi:10.3389 / fpsyg.2013.00667. PMC 3790330. PMID 24109460.
- ^ Robinson, Zack; Maley, Corey J .; Piccinini, Gualtiero (2015). „Je vědomí Spandrel?“. Journal of the American Philosophical Association. 1 (2): 365–383. doi:10.1017 / apa.2014.10.
- ^ Thorpe a kol. 1996
- ^ VanRullen a Koch 2003
- ^ Baars 1988
- ^ Dehaene a kol. 2003
- ^ Koch, Christof (2004). Pátrání po vědomí: neurobiologický přístup. Englewood, US-CO: Roberts & Company Publishers. ISBN 978-0-9747077-0-9.
- ^ Podívejte se na Cooneyho předmluvu k dotisku Chalmersova papíru: Brian Cooney, vyd. (1999). „Kapitola 27: Řešení problému vědomí“. Místo mysli. Cengage Learning. 382 ff. ISBN 978-0534528256.
- ^ Chalmers, David (1995). „Tváří v tvář problému vědomí“. Journal of Consciousness Studies. 2 (3): 200–219. Viz také tento odkaz
Reference
- Adamantidis A.R., Zhang F., Aravanis A.M., Deisseroth K. and de Lecea L. (2007) Neurální substráty probuzení sondované s optogenetickou kontrolou hypocretinových neuronů. Příroda. pokročilá online publikace.
- Baars B.J. (1988) Kognitivní teorie vědomí. Cambridge University Press: New York, NY.
- Sian Beilock, Carr T.H., MacMahon C. a Starkes J.L. (2002) Když je věnování pozornosti kontraproduktivní: dopad pozornosti rozdělené versus dovednosti zaměřené na nováčky a zkušený výkon senzomotorických dovedností. J. Exp. Psychol. Appl. 8: 6–16.
- Blumenfeld H., McNally K.A., Vanderhill S.D., Paige A.L., Chung R., Davis K., Norden A.D., Stokking R., Studholme C., Novotny E.J. Jr., Zubal I.G. and Spencer S.S. (2004) Pozitivní a negativní korelace sítě v epilepsii spánkového laloku. Cereb. Cort. 14: 892–902.
- Bogen J.E. (1995) O neurofyziologii vědomí: I. Přehled. Vědomí a poznání 4: 52–62.
- Chalmers, David J. (Červen 1998), „Co je neurální korelát vědomí?“, In Metzinger, Thomas (vyd.), Neurální korelace vědomí: empirické a koncepční otázky, MIT Press (publikováno září 2000), ISBN 978-0-262-13370-8
- Crick F. a Koch C. (1990) Směrem k neurobiologické teorii vědomí. Semináře z Neuroscience Vol2, 263–275.
- Crick F.C. a Koch C. (1995) Jsme si vědomi nervové aktivity v primární zrakové kůře? Nature 375: 121–3.
- Dehaene S., Sergent C. a Changeux J.P. (2003) Model neuronové sítě spojující subjektivní zprávy a objektivní fyziologické údaje během vědomého vnímání. Proc. Natl. Acad. Sci. USA 100: 8520–5.
- Haynes J.D. a Rees G. (2005) Predikce orientace neviditelných stimulů z činnosti v lidské primární vizuální kůře. Nat. Neurosci. 8: 686–91.
- Kandel Eric R. (2007). Při hledání paměti: Vznik nové vědy o mysli. W. W. Norton & Company. ISBN 978-0393329377.
- Kim CY a Blake R. (2004) Psychofyzikální magie: vykreslování viditelného „neviditelného“. Trendy Cogn. Sci. 9: 381–8.
- Koch, Christof (2004). Pátrání po vědomí: neurobiologický přístup. Englewood, US-CO: Roberts & Company Publishers. ISBN 978-0-9747077-0-9.
- Koch C. a Crick F.C. (2001) O zombie uvnitř. Nature 411: 893.
- Koch C. a Hepp K. (2006) Kvantová mechanika a vyšší funkce mozku: Poučení z kvantového výpočtu a neurobiologie. Nature 440: 611–2. (Volně k dispozici od http://www.theswartzfoundation.org/papers/caltech/koch-hepp-07-final.pdf (2007))
- Kreiman G., Fried I. a Koch C. (2002) Single-neuron koreluje subjektivního vidění v lidském mediálním temporálním laloku. Proc Natl. Acad. Sci. USA 99: 8378–83.
- Laureys S. (2005) Nervový korelát (ne) vědomí: Poučení z vegetativního stavu. Trendy Cogn. Sci. 9: 556–9.
- Lee S.H., Blake R. a Heeger D.J. (2007) Hierarchie kortikálních odpovědí, které jsou základem binokulární rivality. Nat. Neurosci. 10: 1048–54.
- Leopold D.A. a Logothetis N.K. (1996) Změny aktivity v rané zrakové kůře odrážejí vnímání opic během binokulárního soupeření. Nature 379: 549–53.
- Logothetis N. (1998) Jednotlivé jednotky a vědomé vidění. Philos. Trans. R. Soc. Lond. B, 353: 1801–18.
- Milner A.D. a Goodale M.A. (1995) Vizuální mozek v akci. Oxford University Press, Oxford, Velká Británie.
- Owen A.M., Cleman M.R., Boly M., Davis M.H., Laureys S. a Pickard J.D. (2006) Detekce povědomí ve vegetativním stavu. Science 313: 1402.
- Rees G. a Frith C. (2007) Metodiky pro identifikaci nervových korelátů vědomí. In: The Blackwell Companion to Consciousness. Velmans M a Schneider S, eds., Str. 553–66. Blackwell: Oxford, Velká Británie.
- Sheinberg D.L. a Logothetis N.K. (1997) Role temporálních kortikálních oblastí v percepční organizaci. Proc. Natl. Acad. Sci. USA 94: 3408–13.
- Schiff, Nicholas D. (listopad 2004), „Neurologie zhoršeného vědomí: Výzvy pro kognitivní neurovědu.“, V Gazzaniga, Michael S. (vyd.), Kognitivní neurovědy (3. vydání), MIT Press, ISBN 978-0-262-07254-0
- Shimono M. a Niki K. (2013) Global Mapping of the Whole-Brain Network Underlining Binocular Rivalry. Připojení mozku 3: 212-221.
- Thorpe S., Fize D. a Marlot C. (1996) Rychlost zpracování v lidském vizuálním systému. Nature 381: 520–2.
- Squire, Larry R. (2008). Základní neurověda (3. vyd.). Akademický tisk. p.1256. ISBN 978-0-12-374019-9.
- Tononi G. (2004) Teorie informační integrace vědomí. BMC Neuroscience. 5: 42–72.
- Tononi, Giulio a Boly, Melanie a Massimini, Marcello a Koch, Christof (2016). "Integrovaná teorie informací: od vědomí k jeho fyzickému substrátu". Recenze přírody Neurovědy. Nature Publishing Group. 17 (5): 450–461. doi:10.1038 / nrn.2016.44. PMID 27225071.CS1 maint: více jmen: seznam autorů (odkaz)
- VanRullen R. a Koch C. (2003) Vizuální selektivní chování lze vyvolat procesem zpětné vazby. J. Cogn. Neurosci. 15: 209–17.
- Villablanca J.R. (2004) Kontrapunkt funkční role předního mozku a mozkového kmene při kontrole systému probuzení. J. Sleep Res. 13: 179–208.
- Zeman A. (2001) Vědomí. Mozek. 7: 1263-1289.
Další čtení
- Chalmers, D (1995). Vědomá mysl: Hledání základní teorie. Filozofie mysli. Oxford: Oxford University Press. ISBN 9780195117899.
- Dawkins, MS (1993). Pouze našimi očima? Hledání vědomí zvířat. Oxford: Oxford University Press. ISBN 9780198503200.
- Edelman, GM; Tononi, G (2000). Vědomí: Jak se hmota stává představivostí. New York: Základní knihy. ISBN 9780465013777.
- Goodale, MA; Milner, AD (2004). Zrak neviditelný: Zkoumání vědomé a nevědomé vize. Oxford: Oxford University Press. JAKO V B012YXAXS4.
- Koch, Christof (2004). Pátrání po vědomí: neurobiologický přístup. Englewood, US-CO: Roberts & Company Publishers. ISBN 978-0-9747077-0-9.
- Koch C. a Crick F.C. (2001) O zombie uvnitř. Nature 411: 893.
- Koch C. a Hepp K. (2006) Kvantová mechanika a vyšší funkce mozku: Poučení z kvantového výpočtu a neurobiologie. Nature 440: 611–2. (Volně k dispozici od http://www.theswartzfoundation.org/papers/caltech/koch-hepp-07-final.pdf (2007))
- Kreiman G., Fried I. a Koch C. (2002) Single-neuron koreluje subjektivního vidění v lidském mediálním temporálním laloku. Proc Natl. Acad. Sci. USA 99: 8378–83.
- Logothetis, N. K.; Guggenberger, Heinz; Peled, Sharon; Pauls, Jon (1999). "Funkční zobrazování opičího mozku". Přírodní neurovědy. 2 (6): 555–562. doi:10.1038/9210. PMID 10448221.
- Schall, J. "O budování mostu mezi mozkem a chováním "Výroční recenze v psychologii. Sv. 55. únor 2004. str. 23–50.
- Crick, Francis (1994). Úžasná hypotéza: vědecké hledání duše. Macmillan Reference USA. ISBN 978-0-684-19431-8.
- David J. Chalmers (1996). Vědomá mysl: Při hledání základní teorie: Při hledání základní teorie. Oxford University Press, USA. ISBN 978-0-19-511789-9.
- Nikos Logothetis
- Metzinger, T. (vyd.) (2000). „Neurální korelace vědomí“. Cambridge, MA: MIT Press.
- Newsome Lab Publications.
- Atkinson, A. a kol. „Vědomí: Mapování teoretické krajiny“ - Článek o výpočetních korelátech vědomí
- Vaas, Ruediger (1999): „Proč jsou neurální korelace vědomí v pořádku, ale nestačí“. Anthropology & Philosophy Vol. 3, s. 121–141. https://web.archive.org/web/20120205025719/http://www.swif.uniba.it/lei/mind/texts/t0000009.htm