N-vázaná glykosylace - N-linked glycosylation

Nvázaná glykosylace, je připevnění oligosacharid, sacharid skládající se z několika molekul cukru, někdy označovaných také jako glykan k atomu dusíku ( amide dusík an asparagin (Asn) zbytek a protein ), v procesu zvaném N-glykosylace, studoval v biochemie.[1] Tento typ propojení je důležitý pro obě struktury[2] a funkce[3] některých eukaryotických proteinů. The N- propojené glykosylace proces probíhá v eukaryoty a široce v archaea, ale velmi zřídka v bakterie. Povaha Nnavázané glykany připojené k glykoproteinu jsou určeny proteinem a buňkou, ve které jsou exprimovány.[4] Také se liší napříč druh. Různé druhy syntetizují různé typy Nvázaný glykan.
Energetika tvorby vazeb
V glykoproteinu jsou zahrnuty dva typy vazeb: vazby mezi sacharidy zbytky v glykanu a vazba mezi glykanovým řetězcem a molekulou proteinu.
Cukr skupiny jsou navzájem spojeny v glykanovém řetězci prostřednictvím glykosidové vazby. Tyto vazby se obvykle tvoří mezi uhlíky 1 a 4 molekul cukru. Tvorba glykosidové vazby je energeticky nepříznivá, proto je reakce spojena s hydrolýza ze dvou ATP molekuly.[4]
Na druhou stranu připojení glykanového zbytku k proteinu vyžaduje rozpoznání a konsensuální sekvence. Nvázané glykany jsou téměř vždy připojeny k dusík atom asparaginového (Asn) postranního řetězce, který je přítomen jako součást Asn – X–Ser /Thr konsensuální sekvence, kde X je jakákoli aminokyselina kromě prolin (Pro).[4]
V živočišných buňkách je glykan navázaný na asparagin téměř nevyhnutelně N-acetylglukosamin (GlcNAc) v β-konfiguraci.[4] Tato p-vazba je podobná glykosidové vazbě mezi cukrovými skupinami ve struktuře glykanu, jak je popsáno výše. Místo toho, aby byl připoután k cukru hydroxyl skupina, anomerní uhlík atom je připojen k amidovému dusíku. Energie potřebná pro toto propojení pochází z hydrolýza a pyrofosfát molekula.[4]
Biosyntéza

Biosyntéza N- vázané glykany probíhají ve 3 hlavních krocích:[4]
- Syntéza dolicholem vázaného prekurzorového oligosacharidu
- En bloc přenos prekurzorového oligosacharidu na protein
- Zpracování oligosacharidu
Syntéza, přenos en bloc a počáteční oříznutí prekurzoru oligosacharid se vyskytuje v endoplazmatické retikulum (ER). Následné zpracování a modifikace oligosacharidového řetězce se provádí v Golgiho aparát.
Syntéza glykoproteinů je tak prostorově oddělena v různých buněčných kompartmentech. Proto typ Nsyntetizovaný glykan, závisí na jeho dostupnosti pro různé enzymy přítomné v těchto buněčných kompartmentech.
I přes rozmanitost však vše N-glykany jsou syntetizovány společnou cestou se společnou základní glykanovou strukturou.[4]Struktura jádra glykanu je v podstatě tvořena dvěma N-acetylglukosamin a tři manóza zbytky. Tento jádrový glykan je poté dále zpracován a upraven, což má za následek pestrou škálu N-glykanové struktury.[4]
Syntéza prekurzorového oligosacharidu
Proces Nnavázaná glykosylace začíná tvorbou dolichol vázaný cukr GlcNAc. Dolichol je a lipid molekula složená z opakování isopren Jednotky. Tato molekula se nalézá připojená k membráně ER. Molekuly cukru jsou připojeny k dolicholu prostřednictvím pyrofosfátové vazby[4] (jeden fosfát byl původně spojen s dolicholem a druhý fosfát pocházel z nukleotid cukr). Oligosacharidový řetězec se poté prodlužuje přidáváním různých molekul cukru postupným způsobem za vzniku prekurzorového oligosacharidu.
Sestavení tohoto prekurzorového oligosacharidu probíhá ve dvou fázích: fáze I a II.[4] Fáze I se odehrává na cytoplazmatický strana ER a fáze II se odehrává na luminální straně ER.
Prekurzorová molekula připravená k přenosu na protein se skládá ze 2 GlcNAc, 9 manózy a 3 glukóza molekuly.

| |
| Fáze II | |
je dárcem rezidua manózy (formace: Dol-P + GDP-Man → Dol-P-Man + GDP) a Dol-P-Gluc je dárcem rezidua glukózy (formace: Dol-P + UDP-Glc → Dol-P- Glc + UDP).
| |
Přenos glykanu na protein
Jakmile se vytvoří prekurzorový oligosacharid, dokončený glykan se poté přenese do rodícího se organismu polypeptid v lumen membrány ER. Tato reakce je řízena energií uvolněnou ze štěpení pyrofosfátové vazby mezi molekulou dolichol-glykanu. Před přenosem glykanu na rodící se polypeptid je třeba splnit tři podmínky:[4]
- Asparagin musí být umístěn v konkrétní shodné sekvenci v primární struktura (Asn – X – Ser nebo Asn – X – Thr nebo ve vzácných případech Asn – X – Cys).[5]
- Asparagin musí být vhodně umístěn v trojrozměrné struktuře proteinu (cukry jsou polární molekuly a proto musí být navázán na asparagin nacházející se na povrchu proteinu a nesmí být pohřben v proteinu)
- Asparagin musí být nalezen v luminální straně endoplazmatického retikula pro N-spojená glykosylace. Cílové zbytky se nacházejí buď v sekrečních proteinech, nebo v oblastech transmembránový protein který stojí před lumen.
Oligosacharyltransferáza je enzym odpovědný za rozpoznání konsensuální sekvence a přenos prekurzorového glykanu na akceptor polypeptidu, který je translatován v lumenu endoplazmatického retikula. N-vázaná glykosylace je tedy společná translační událost
Zpracování glykanu

N- zpracování glykanů se provádí v endoplazmatickém retikulu a Golgiho těle. K počátečnímu ořezávání molekuly prekurzoru dochází v ER a následné zpracování probíhá v Golgi.
Po přenosu dokončeného glykanu na rodící se polypeptid jsou ze struktury odstraněny dva zbytky glukózy. Enzymy známé jako glykosidázy odstraňují některé zbytky cukru. Tyto enzymy mohou pomocí molekuly vody rozbít glykosidické vazby. Tyto enzymy jsou exoglykosidázy, protože pouze fungují monosacharid zbytky umístěné na neredukujícím konci glykanu.[4] Tento počáteční krok ořezávání se považuje za krok kontroly kvality v ER, který se má monitorovat skládání bílkovin.
Jakmile je protein správně složen, jsou odstraněny dva zbytky glukózy glukosidáza I a II. Odstranění posledního třetího zbytku glukózy signalizuje, že glykoprotein je připraven k přenosu z ER do cis-Golgi.[4] . ER manosidáza katalyzuje odstranění této konečné glukózy. Pokud však protein není správně složen, nejsou odstraněny zbytky glukózy, a tak glykoprotein nemůže opustit endoplazmatické retikulum. A garde bílkoviny (kalnexin /kalretikulin ) váže se na rozložený nebo částečně složený protein, aby pomohl skládání proteinu.
Další krok zahrnuje další přidání a odstranění zbytků cukru v cis-Golgi. Tyto modifikace jsou katalyzovány glykosyltransferázami a glykosidázami. V cis-Golgi, řada mannosidáz odstraňuje některé nebo všechny čtyři manosové zbytky ve vazbách α-1,2.[4] Zatímco v mediální části Golgiho glykosyltransferázy přidávají zbytky cukru ke struktuře jádra glykanu, čímž vznikají tři hlavní typy glykanů: vysoký obsah manózy, hybridní a komplexní glykany.
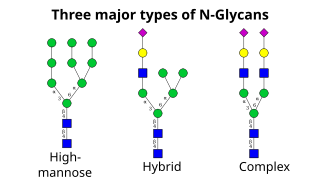
- High-mannose jsou v podstatě jen dva N-acetylglukosaminy s mnoha zbytky manózy, často téměř tolik, kolik je pozorováno v prekurzorových oligosacharidech před jeho připojením k proteinu.
- Komplexní oligosacharidy jsou tak pojmenovány, protože mohou obsahovat téměř jakýkoli počet dalších typů sacharidů, včetně více než původních dvou N-acetylglukosaminy.
- Hybridní oligosacharidy obsahují manosové zbytky na jedné straně větve, zatímco na druhé straně a N-acetylglukosamin iniciuje komplexní větev.
Pořadí přidávání cukrů k rostoucím glykanovým řetězcům je určeno specifičností substrátů enzymů a jejich přístupem k substrátu při jejich pohybu sekreční cesta. Organizace tohoto aparátu v buňce tedy hraje důležitou roli při určování, které glykany se vyrábějí.
Enzymy v Golgi
Golgiho enzymy hrají klíčovou roli při určování syntézy různých typů glykanů. Pořadí působení enzymů se odráží v jejich poloze v Golgiho zásobníku:
| Enzymy | Místo v Golgi |
|---|---|
| Mannosidáza I | cis-Golgi |
| GlcNAc transferázy | mediální Golgi |
| Galaktosyltransferáza a Sialyltransferáza | trans-Golgi |
U archaea a prokaryot
Podobný NCesta biosyntézy glykanu byla nalezena u prokaryot a Archea.[6] Ve srovnání s eukaryoty se však nezdá, že by se konečná glykánová struktura v eubakteriích a archaeách příliš lišila od počátečního prekurzoru vytvořeného v endoplazmatickém retikulu. U eukaryot je původní prekurzorový oligosacharid značně modifikován na cestě k buněčnému povrchu.[4]
Funkce
Nvázané glykany mají vnitřní a vnější funkce.[4]
V rámci imunitního systému Nnavázané glykany na povrchu imunitní buňky pomohou diktovat migrační vzorec buňky, např. imunitní buňky, které migrují do kůže, mají specifické glykosylace, které upřednostňují navádění na dané místo.[7] Glykosylační vzorce na různých imunoglobulinech včetně IgE, IgM, IgD, IgA a IgG jim propůjčují jedinečné efektorové funkce tím, že mění jejich afinitu k Fc a dalším imunitním receptorům.[7] Glykany mohou být také zapojeny do diskriminace „já“ a „ne sebe“, což může být relevantní pro patofyziologii různých autoimunitních onemocnění.[7]
| Vnitřní |
|
| Vnější |
|
Klinický význam
Změny v Nnavázaná glykosylace byla spojena s různými nemocemi včetně revmatoidní artritida,[9] cukrovka 1. typu,[10] Crohnova nemoc,[11] a rakoviny.[12][13]
Mutace v osmnácti genech zapojených do Nnavázaná glykosylace vede k řadě onemocnění, z nichž většina zahrnuje nervový systém.[3][13]
Význam terapeutických proteinů
Mnoho terapeutický bílkoviny na trhu jsou protilátky, což jsou Nspojené glykoproteiny. Například, Etanercept, Infliximab a Rituximab jsou N-glykosylované terapeutické proteiny.

Důležitost Nglykosylace spojená s vazbami je stále více zřejmá v oboru léčiva.[14] Ačkoli bakteriální nebo kvasinky produkce bílkovin systémy mají významné potenciální výhody, jako je vysoký výtěžek a nízké náklady, nastávají problémy, když je zájmovým proteinem glykoprotein. Většina prokaryotických expresních systémů, jako je E-coli nemůže provést posttranslační úpravy. Na druhé straně, eukaryotičtí expresní hostitelé, jako jsou kvasinky a zvířecí buňky, mají různé glykosylační vzorce. Proteiny produkované v těchto expresních hostitelích často nejsou totožné s lidským proteinem, a proto jsou příčinou imunogenní reakce u pacientů. Například, S. cerevisiae (kvasinky) často produkují vysoce manosové glykany, které jsou imunogenní.
Nehumánní savčí expresní systémy, jako jsou CHO nebo Buňky NS0 mít potřebné vybavení pro přidávání komplexních glykanů lidského typu. Glykany produkované v těchto systémech se však mohou lišit od glykanů produkovaných u lidí, protože mohou být uzavřeny oběma N-glykolylneuraminová kyselina (Neu5Gc) a N-acetylneuraminová kyselina (Neu5Ac), zatímco lidské buňky produkují pouze glykoproteiny obsahující N-acetylneuraminová kyselina. Kromě toho mohou zvířecí buňky také produkovat glykoproteiny obsahující galaktóza-alfa-1,3-galaktóza epitop, který může vyvolat závažné alergenní reakce, včetně anafylaktický šok u lidí, kteří mají Alfa-gal alergie.
Tyto nevýhody byly vyřešeny několika přístupy, jako je eliminace drah, které produkují tyto glykanové struktury prostřednictvím genetických knockoutů. Kromě toho byly geneticky upraveny další expresní systémy k produkci terapeutických glykoproteinů podobných člověku Nvázané glykany. Patří mezi ně kvasinky jako Pichia pastoris,[15] buněčné linie hmyzu, zelené rostliny,[16] a dokonce i bakterie.
Viz také
Reference
- ^ "Glykosylace". UniProt: Sekvence proteinů a funkční informace.
- ^ Imperiali B, O'Connor SE (prosinec 1999). "Účinek N-vázaná glykosylace na glykopeptidové a glykoproteinové struktuře ". Aktuální názor na chemickou biologii. 3 (6): 643–9. doi:10.1016 / S1367-5931 (99) 00021-6. PMID 10600722.
- ^ A b Patterson MC (září 2005). "Metabolické napodobeniny: poruchy N-vázaná glykosylace ". Semináře z dětské neurologie. 12 (3): 144–51. doi:10.1016 / j.spen.2005.10.002. PMID 16584073.
- ^ A b C d E F G h i j k l m n Ó p Drickamer K, Taylor ME (2006). Úvod do glykobiologie (2. vyd.). Oxford University Press, USA. ISBN 978-0-19-928278-4.
- ^ Mellquist JL, Kasturi L, Spitalnik SL, Shakin-Eshleman SH (květen 1998). „Aminokyselina následující po sekvenci Asn – X – Ser / Thr je důležitým determinantem N- účinnost glykosylace spojeného jádra ". Biochemie. 37 (19): 6833–7. doi:10.1021 / bi972217k. PMID 9578569.
- ^ Dell A, Galadari A, Sastre F, Hitchen P (2010). „Podobnosti a rozdíly v mechanismech glykosylace u prokaryot a eukaryot“. International Journal of Microbiology. 2010: 1–14. doi:10.1155/2010/148178. PMC 3068309. PMID 21490701.
- ^ A b C Maverakis E, Kim K, Shimoda M, Gershwin ME, Patel F, Wilken R a kol. (Únor 2015). „Glykany v imunitním systému a The Altered Glycan Theory of Autoimmunity: a critical review“. Časopis autoimunity. 57 (6): 1–13. doi:10.1016 / j.jaut.2014.12.002. PMC 4340844. PMID 25578468.
- ^ Sinclair AM, Elliott S (srpen 2005). „Glykoinženýrství: účinek glykosylace na vlastnosti terapeutických proteinů“. Journal of Pharmaceutical Sciences. 94 (8): 1626–35. doi:10.1002 / jps.20319. PMID 15959882.
- ^ Nakagawa H, Hato M, Takegawa Y, Deguchi K, Ito H, Takahata M a kol. (Červen 2007). „Detekce změněných N-glykanové profily v celém séru pacientů s revmatoidní artritidou ". Journal of Chromatography B. 853 (1–2): 133–7. doi:10.1016 / j.jchromb.2007.03.003. hdl:2115/28276. PMID 17392038.
- ^ Bermingham ML, Colombo M, McGurnaghan SJ, Blackbourn LA, Vučković F, Pučić Baković M a kol. (Leden 2018). "N-Glykanový profil a onemocnění ledvin u diabetu 1. typu ". Péče o cukrovku. 41 (1): 79–87. doi:10.2337 / dc17-1042. PMID 29146600.
- ^ Trbojević Akmačić I, Ventham NT, Theodoratou E, Vučković F, Kennedy NA, Krištić J a kol. (Červen 2015). „Zánětlivé onemocnění střev souvisí s prozánětlivým potenciálem imunoglobulinu G glycome“. Zánětlivá onemocnění střev. 21 (6): 1237–47. doi:10.1097 / MIB.0000000000000372. PMC 4450892. PMID 25895110.
- ^ Kodar K, Stadlmann J, Klaamas K, Sergeyev B, Kurtenkov O (leden 2012). „Imunoglobulin G Fc Nprofil glykanu u pacientů s rakovinou žaludku metodou LC-ESI-MS: vztah k progresi a přežití nádoru “. Glycoconjugate Journal. 29 (1): 57–66. doi:10.1007 / s10719-011-9364-z. PMID 22179780.
- ^ A b Chen G, Wang Y, Qin X, Li H, Guo Y, Wang Y a kol. (Srpen 2013). "Změna IgG1 Fc Nsouvisející glykosylace u lidské rakoviny plic: diagnostický potenciál související s věkem a pohlavím ". Elektroforéza. 34 (16): 2407–16. doi:10.1002 / elps.201200455. PMID 23766031.
- ^ Dalziel M, Crispin M, Scanlan CN, Zitzmann N, Dwek RA (leden 2014). "Nové zásady pro terapeutické využití glykosylace". Věda. 343 (6166): 1235681. doi:10.1126 / science.1235681. PMID 24385630.
- ^ Hamilton SR, Bobrowicz P, Bobrowicz B, Davidson RC, Li H, Mitchell T a kol. (Srpen 2003). "Produkce komplexních lidských glykoproteinů v kvasnicích". Věda. 301 (5637): 1244–6. doi:10.1126 / science.1088166. PMID 12947202.
- ^ Strasser R, Altmann F, Steinkellner H (prosinec 2014). "Řízená glykosylace rostlinných produkovaných rekombinantních proteinů". Aktuální názor na biotechnologie. 30: 95–100. doi:10.1016 / j.copbio.2014.06.008. PMID 25000187.
externí odkazy
- GlycoEP: In silico Platforma pro předpověď N-, Ó- a C-Glykosity v eukaryotických proteinových sekvencích
- Maverakis E, Kim K, Shimoda M, Gershwin ME, Patel F, Wilken R, Raychaudhuri S, Ruhaak LR, Lebrilla CB (únor 2015). „Glykany v imunitním systému a The Altered Glycan Theory of Autoimmunity: a critical review“. Časopis autoimunity. 57: 1–13. doi:10.1016 / j.jaut.2014.12.002. PMC 4340844. PMID 25578468.