Esenciální gen - Essential gene
Esenciální geny jsou nepostradatelné geny pro organismy k růstu a reprodukci potomků v určitém prostředí.[1] Nicméně, být nezbytný je vysoce závislá na okolnostech, ve kterých organismus žije. Například gen potřebný k trávení škrob je nezbytné pouze tehdy, je-li škrob jediným zdrojem energie. Nedávno byly učiněny systematické pokusy identifikovat ty geny, které jsou naprosto nezbytné pro udržení života, za předpokladu, že jsou k dispozici všechny živiny.[2] Takové experimenty vedly k závěru, že absolutně požadovaný počet genů pro bakterie je řádově kolem 250–300. Esenciální geny jednobuněčných organismů kódují proteiny pro tři základní funkce, včetně zpracování genetické informace, buněčných obalů a výroby energie.[1] Tyto genové funkce se používají k udržení centrální polohy metabolismus, replikovat DNA, přeložit geny do bílkoviny, udržují základní buněčnou strukturu a zprostředkovávají transportní procesy do a z buňky. Většina genů není nezbytná, ale vyjadřuje se selektivní výhody a zvýšil zdatnost. Ve srovnání s jednobuněčnými organismy mají mnohobuněčné organismy podstatnější geny související s komunikací a vývojem. Většina základních genů ve virech souvisí se zpracováním a udržováním genetické informace. Na rozdíl od většiny jednobuněčných organismů chybí virům mnoho základních genů pro metabolismus,[1] což je nutí unést metabolismus hostitele. Většina genů není nezbytná, ale vyjadřuje se selektivní výhody a zvýšil zdatnost. Drtivá většina genů proto není nezbytná a mnoho z nich může být odstraněno bez následků, alespoň za většiny okolností.
Bakterie: studie na celém genomu
K identifikaci základních genů na celém genomu byly použity dvě hlavní strategie: řízená delece genů a náhodná mutageneze použitím transpozice. V prvním případě anotované jednotlivé geny (nebo ORF ) jsou zcela odstraněny z genom systematicky. Při mutagenezi zprostředkované transposony jsou transpozony náhodně vloženy na co nejvíce pozic v genomu, jejichž cílem je narušit funkci cílených genů (viz obrázek níže). Inzerční mutanti, kteří jsou stále schopni přežít nebo růst, naznačují transposon vložený do genu, který není nezbytný pro přežití. Umístění inzercí transposonu lze určit hybridizací na mikročipy [3] nebo skrz sekvenování transpozonů . Souhrn těchto obrazovek je uveden v tabulce.[2][4]
| Organismus | Mutageneze | Metoda | Odečet | ORF | Non-ess. | Nezbytný | % Ess. | Poznámky | Čj. |
|---|---|---|---|---|---|---|---|---|---|
| Mycoplasma genitalium / pneumoniae | Náhodný | Populace | Sekvenování | 482 | 130 | 265–350 | 55–73% | --- | [5] |
| Mycoplasma genitalium | Náhodný | Klony | Sekvenování | 482 | 100 | 382 | 79% | před naším letopočtem | [6] |
| Zlatý stafylokok WCUH29 | Náhodný | Klony | Sekvenování | 2,600 | n / a | 168 | n / a | před naším letopočtem | [7] |
| Zlatý stafylokok RN4220 | Náhodný | Klony | Sekvenování | 2,892 | n / a | 658 | 23% | --- | [8] |
| Haemophilus influenzae Rd | Náhodný | Populace | Footprint-PCR | 1,657 | 602 | 670 | 40% | --- | [9] |
| Streptococcus pneumoniae Rx-1 | Cílené | Klony | Tvorba kolonií | 2,043 | 234 | 113 | n / a | C | [10] |
| Streptococcus pneumoniae D39 | Cílené | Klony | Tvorba kolonií | 2,043 | 560 | 133 | n / a | C | [11] |
| Streptococcus pyogenes 5448 | Náhodný | Transposon | Tn-seq | 1,865 | ? | 227 | 12% | --- | [12] |
| Streptococcus pyogenes NZ131 | Náhodný | Transposon | Tn-seq | 1,700 | ? | 241 | 14% | --- | [12] |
| Streptococcus sanguinis SK36 | Cílené | Klony | Tvorba kolonií | 2,270 | 2,052 | 218 | 10% | aj | [1][13] |
| Mycobacterium tuberculosis H37Rv | Náhodný | Populace | Microarray | 3,989 | 2,567 | 614 | 15% | --- | [14] |
| Mycobacterium tuberculosis | Náhodný | Transposon | ? | 3,989 | ? | 401 | 10% | --- | [15] |
| Mycobacterium tuberculosis H37Rv | Náhodný | Transposon | Sekvenování NG | 3,989 | ? | 774 | 19% | --- | [16][17] |
| Mycobacterium tuberculosis H37Rv | Náhodný | Transposon | Sekvenování NG | 3,989 | 3,364 | 625 | 16% | Ahoj | [18] |
| Mycobacterium tuberculosis | --- | Výpočetní | Výpočetní | 3,989 | ? | 283 | 7% | --- | [19] |
| Bacillus subtilis 168 | Cílené | Klony | Tvorba kolonií | 4,105 | 3,830 | 261 | 7% | a, d, g | [20][21] |
| Escherichia coli K-12 MG1655 | Náhodný | Populace | Footprint-PCR | 4,308 | 3,126 | 620 | 14% | --- | [22] |
| Escherichia coli K-12 MG1655 | Cílené | Klony | Tvorba kolonií | 4,308 | 2,001 | n / a | n / a | a, e | [23] |
| Escherichia coli K-12 BW25113 | Cílené | Klony | Tvorba kolonií | 4,390 | 3,985 | 303 | 7% | A | [24] |
| Pseudomonas aeruginosa PAO1 | Náhodný | Klony | Sekvenování | 5,570 | 4,783 | 678 | 12% | A | [25] |
| Porphyromonas gingivalis | Náhodný | Transposon | Sekvenování | 1,990 | 1,527 | 463 | 23% | --- | [26] |
| Pseudomonas aeruginosa PA14 | Náhodný | Klony | Sekvenování | 5,688 | 4,469 | 335 | 6% | a, f | [27] |
| Salmonella typhimurium | Náhodný | Klony | Sekvenování | 4,425 | n / a | 257 | ~11% | před naším letopočtem | [28] |
| Helicobacter pylori G27 | Náhodný | Populace | Microarray | 1,576 | 1,178 | 344 | 22% | --- | [29] |
| Corynebacterium glutamicum | Náhodný | Populace | ? | 3,002 | 2,352 | 650 | 22% | --- | [30] |
| Francisella novicida | Náhodný | Transposon | ? | 1,719 | 1,327 | 392 | 23% | --- | [31] |
| Mycoplasma pulmonis UAB CTIP | Náhodný | Transposon | ? | 782 | 472 | 310 | 40% | --- | [34] |
| Vibrio cholerae N16961 | Náhodný | Transposon | ? | 3,890 | ? | 779 | 20% | --- | [35] |
| Salmonella Typhi | Náhodný | Transposon | ? | 4,646 | ? | 353 | 8% | --- | [36] |
| Zlatý stafylokok | Náhodný | Transposon | ? | ~2,600 | ? | 351 | 14% | --- | [37] |
| Caulobacter crescentus | Náhodný | Transposon | Tn-Seq | 3,876 | 3,240 | 480 | 12.2% | --- | [38] |
| Neisseria meningitidis | Náhodný | Transposon | ? | 2,158 | ? | 585 | 27% | --- | [39] |
| Desulfovibrio alaskensis | Náhodný | Transposon | Sekvenování | 3,258 | 2,871 | 387 | 12% | --- | [40] |
Tabulka 1. Esenciální geny v bakteriích. Mutageneze: cílené mutanty jsou genové delece; náhodný mutanti jsou transposon vložení. Metody: Klony označují delece jednoho genu, populace označuje mutagenezi celé populace, např. pomocí transpozonů. Základní geny z populačních obrazovek zahrnují geny nezbytné pro fitness (viz text). ORF: počet všech otevřené čtecí rámce v tom genomu. Poznámky: a) je k dispozici sbírka mutantů; (b) metoda přímého screeningu esenciálnosti (např. prostřednictvím antisense RNA), která neposkytuje informace o neesenciálních genech. (c) K dispozici je pouze částečný soubor údajů. (d) Zahrnuje předpokládanou genovou esenciálnost a kompilaci dat ze zveřejněných studií esenciální aktivity jednoho genu. e) Probíhá projekt. f) Sníženo porovnáním dvou datových souborů genové esence získaných nezávisle v P. aeruginosa kmeny PA14 a PAO1. (g) Původní výsledek 271 esenciálních genů byl opraven na 261, přičemž 31 genů, které byly považovány za esenciální, bylo ve skutečnosti nepodstatné, zatímco od té doby bylo popsáno 20 nových esenciálních genů.[21] (h) Počítání genů se základními doménami a těch, které vedou k poruchám růstu, pokud jsou narušeny jako zásadní, a těch, které vedou k růstovým výhodám, pokud jsou narušeny jako nepodstatné. (i) Podílela se na plně nasycené mutantní knihovně 14 replikátů s 84,3% možných inzertních míst s alespoň jednou transpozonovou inzercí. (j) Každý základní gen byl nezávisle potvrzen nejméně pětkrát.
Na základě genomových experimentů a analýz systémové biologie byla vyvinuta základní genová databáze Kong a kol. (2019) pro předpovídání> 4000 bakteriálních druhů.[41]
Eukaryoty
v Saccharomyces cerevisiae (nadějné kvasinky) 15–20% všech genů je nezbytných. v Schizosaccharomyces pombe (štěpné kvasinky) bylo zkonstruováno 4 836 heterozygotních delecí pokrývajících 98,4% z 4 914 proteinů kódujících otevřené čtecí rámce. Ukázalo se, že 1260 z těchto výmazů bylo nezbytných.[42]
Podobné obrazovky je obtížnější provádět v jiných mnohobuněčných organismech, včetně savci (jako model pro člověka), z technických důvodů, a jejich výsledky jsou méně jasné. Pro červ hlístice však byly vyvinuty různé metody C. elegans,[43] ovocná muška,[44] a zebrafish[45] (viz tabulka). Nedávná studie s 900 myšími geny dospěla k závěru, že 42% z nich bylo esenciálních, i když vybrané geny nebyly reprezentativní.[46]
Pokusy s genovým knockoutem nejsou u lidí možné nebo přinejmenším nejsou etické. Přirozené mutace však vedly k identifikaci mutací, které vedou k časné embryonální nebo pozdější smrti.[47] Všimněte si, že mnoho genů u lidí není absolutně nezbytných pro přežití, ale při mutaci může způsobit závažné onemocnění. Tyto mutace jsou katalogizovány v Online Mendelian Inheritance in Man (OMIM) databáze. Ve výpočetní analýze genetických variací a mutací u 2 472 člověk ortology známých esenciálních genů u myší, Georgi et al. našel silnou, čistící selekci a srovnatelně snížené úrovně variací sekvencí, což naznačuje, že i tyto lidské geny jsou nezbytné.[48]
I když může být obtížné prokázat, že gen je u lidí nezbytný, lze prokázat, že gen je ne zásadní nebo dokonce nezpůsobující nemoc. Například sekvenování genomů 2 636 islandských občanů a genotypizace 101 584 dalších subjektů našlo 8 041 jedinců, kteří měli 1 gen zcela vyřazený (tj. Tito lidé byli homozygotní pro nefunkční gen).[49] Z 8 041 jedinců s úplným vyloučením bylo odhadováno 6 885 osob homozygoti Bylo odhadnuto, že 1,249 je sloučenina heterozygoti (tj. měli oba alely genu vyřazeného, ale obě alely měly různé mutace). U těchto jedinců bylo celkem 1171 z 19 135 lidí (RefSeq ) geny (6,1%) byly zcela vyřazeny. Byl vyvozen závěr, že těchto 1171 genů je nepodstatné u lidí - přinejmenším nebyly hlášeny žádné související choroby.[49] Podobně exome sekvence 3222 dospělých pákistánských dědiců s vysokou příbuzností rodičů odhalily 1111 homozygotních genotypů vzácných variant s předpokládanou ztrátou genové funkce (LOF = knockouts) v 781 genech.[50] Tato studie zjistila v průměru 140 předpokládaných genotypů LOF (na subjekt), včetně 16 vzácných (méně významných) alela frekvence <1%) heterozygotů, 0,34 vzácných homozygotů, 83,2 běžných heterozygotů a 40,6 společných homozygotů. Téměř všechny vzácné homozygotní genotypy LOF byly nalezeny uvnitř autozygotní segmenty (94,9%).[50] I když většina z těchto jedinců neměla žádné zjevné zdravotní problémy vyplývající z jejich vadných genů, je možné, že při podrobnějším zkoumání lze nalézt menší zdravotní problémy.
Souhrn obrazovek nezbytnosti je uveden v tabulce níže (většinou na základě databáze základních genů.[51]
| Organismus | Metoda | Esenciální geny | Čj. |
| Arabidopsis thaliana | Inzerce T-DNA | 777 | [52] |
| Caenorhabditis elegans (červ) | Interference RNA | 294 | [43] |
| Danio rerio (zebrafish) | Inzerční mutageneze | 288 | [45] |
| Drosophila melanogaster (ovocný let) | Mutageneze inzerce P-prvku | 339 | [44] |
| Homo sapiens (člověk) | Hledání literatury | 118 | [47] |
| Homo sapiens (člověk) | Obrazovka založená na CRISPR / Cas9 | 1,878 | [53] |
| Homo sapiens (člověk) | Haploidní obrazovka pro zachycení genů | ~2,000 | [54] |
| Homo sapiens (člověk) | myší ortology | 2,472 | [55] |
| Mus musculus (myš) | Hledání literatury | 2114 | [56] |
| Saccharomyces cerevisiae (droždí) | Delece jednoho genu | 878 | [57] |
| Saccharomyces cerevisiae (droždí) | Delece jednoho genu | 1,105 | [58] |
| Schizosaccharomyces pombe (droždí) | Delece jednoho genu | 1,260 | [42] |
Viry
Virům chybí mnoho genů nezbytných pro metabolismus,[1] nutit je k únosu metabolismu hostitele. Screening esenciálních genů byl proveden u několika virů. Například, lidský cytomegalovirus (CMV) bylo zjištěno, že má 41 základních, 88 nepodstatných a 27 rozšiřujících ORF (celkem 150 ORF). Nejpodstatnější a rozšiřující geny se nacházejí v centrální oblasti a neesenciální geny se obecně shlukují blízko konců virového genomu.[59]
Tscharke a Dobson (2015) sestavili komplexní průzkum základních genů v roce Virus vakcínie a přidělili role každému z 223 ORF kmene Western Reserve (WR) a 207 ORF kodaňského kmene, přičemž se vyhodnotila jejich role v replikaci v buněčné kultuře. Podle jejich definice je gen považován za nezbytný (tj. Má roli v buněčné kultuře), pokud jeho delece vede ke snížení titru viru více než 10krát v jednokrokové nebo vícestupňové růstové křivce. Všechny geny zapojené do produkce zabaleného virionu, tvorby aktinového ocasu a uvolňování extracelulárního virionu byly také považovány za nezbytné. Geny, které ovlivňují velikost plaku, ale ne replikaci, byly definovány jako nepodstatné. Podle této definice je pro replikaci viru vakcínie požadováno 93 genů v buněčné kultuře, zatímco 108 a 94 ORF z WR a Kodaně není nezbytných.[60] Viry vakcínie s delecemi na obou koncích genomu se chovaly podle očekávání a vykazovaly pouze mírné defekty nebo defekty hostitele. Naproti tomu kombinace delecí na obou koncích genomu pro kmen VACV WR způsobila devastující růstový defekt na všech testovaných buněčných liniích. To ukazuje, že delece jednoho genu nejsou dostatečné k posouzení esenciality genů a že ve viru Vaccinia je esenciálních více genů, než se původně myslelo.[60]
Jeden z bakteriofágy testováno na základní geny zahrnuje mykobakteriofág Giles. Nejméně 35 ze 78 předpovězených genů Giles (45%) není pro lytický růst nezbytných. Bylo zjištěno, že 20 genů je nezbytných.[61] Hlavním problémem fágových genů je, že většina jejich genů zůstává funkčně neznámá, a proto je obtížné posoudit jejich roli. Obrazovka Salmonella enterica fág SPN3US odhalil 13 základních genů, i když zůstává trochu nejasné, kolik genů bylo skutečně testováno.[62]
Kvantitativní analýza genové esence
Teoreticky jsou základní geny kvalitativní.[1] V závislosti na okolním prostředí však mohou určité základní genové mutanty vykazovat dílčí funkce, které lze v některých studiích kvantitativně stanovit. Například konkrétní delece genu může snížit rychlost růstu (nebo míru plodnosti nebo jiné znaky) na 90% divokého typu. Pokud existují isozymy nebo alternativní cesty pro základní geny, mohou být zcela odstraněny.[1]
Syntetická letalita
Dva geny jsou syntetické letální, pokud ani jeden není nezbytný, ale když jsou mutovány oba, dvojitý mutant je smrtelný. Některé studie odhadují, že počet syntetických letálních genů může být řádově 45% všech genů.[63][64]
Podmíněně esenciální geny

Mnoho genů je nezbytných pouze za určitých okolností. Například pokud jde o aminokyselinu lysin je dodáván do buňky jakýkoli gen, který je vyžadován pro výrobu lysinu, není nezbytný. Pokud však není dodáván žádný lysin, stávají se nezbytné geny kódující enzymy pro biosyntézu lysinu, protože bez lysinu není možná žádná syntéza proteinů.[4]
Streptococcus pneumoniae Zdá se, že vyžaduje 147 genů pro růst a přežití v sliny,[65] více než 113–133, které byly nalezeny v předchozích studiích.
Může dojít k deleci genu smrt nebo v bloku buněčné dělení. Zatímco druhý případ může po určitou dobu znamenat „přežití“, bez buněčného dělení může buňka nakonec nakonec zemřít. Podobně se místo blokovaného buněčného dělení mohla buňka zmenšit růst nebo metabolismus od téměř nezjistitelného až po téměř normální. Existuje tedy přechod od „základního“ k zcela nepodstatnému, opět v závislosti na stavu. Někteří autoři tak rozlišovali mezi geny “nezbytné pro přežití" a "nezbytné pro fitness".[4]
Role genetického pozadí. Podobně jako podmínky prostředí může genetické pozadí určit podstatnost genu: gen může být esenciální u jednoho jedince, ale ne u jiného, vzhledem k jeho genetickému pozadí. Genové duplikace jsou jedním z možných vysvětlení (viz níže).
Metabolická závislost. Geny podílející se na určitých biosyntetických drahách, jako např syntéza aminokyselin, se mohou stát neesenciálními, pokud je jedna nebo více aminokyselin dodáno kultivačním médiem[1] nebo jiným organismem.[66] To je hlavní důvod, proč mnoho parazitů (např. Cryptosporidium hominis )[67] nebo endosymbiontové bakterie ztratil mnoho genů (např. Chlamydie ). Takové geny mohou být nezbytné, ale pouze v hostitelském organismu. Například, Chlamydia trachomatis nemůže syntetizovat purin a pyrimidin nukleotidy de novo, takže tyto bakterie jsou závislé na nukleotidových biosyntetických genech hostitele.[68]
Genové duplikace a alternativní metabolické cesty
Mnoho genů je v genomu duplikováno a mnoho organismů má různé metabolické dráhy (alternativní metabolická cesta)[1]) k syntéze stejných produktů. Takový duplikace (paralogy ) a alternativní metabolické cesty často činí esenciální geny neesenciálními, protože duplikát může nahradit původní kopii. Například gen kódující enzym aspartokináza je zásadní v E-coli. Naproti tomu Bacillus subtilis genom obsahuje tři kopie tohoto genu, z nichž žádný není sám o sobě nezbytný. Trojnásobná delece všech tří genů je však smrtelná. V takových případech lze esenciálnost genu nebo skupiny paralogů často předpovědět na základě esenciality esenciálního jediného genu u různých druhů. V kvasinkách je v genomu duplikováno několik esenciálních genů: 8,5% neesenciálních genů, ale pouze 1% esenciálních genů má homolog v genomu kvasinek.[58]
V červi C. elegans, neesenciální geny jsou mezi duplikáty vysoce nadměrně zastoupeny, pravděpodobně proto, že duplikace esenciálních genů způsobuje nadměrnou expresi těchto genů. Woods a kol. zjistili, že neesenciální geny jsou častěji úspěšně duplikovány (fixovány) a ztraceny ve srovnání s esenciálními geny. Naproti tomu základní geny jsou méně často duplikovány, ale po úspěšné duplikaci jsou udržovány po delší dobu.[69]
Zachování
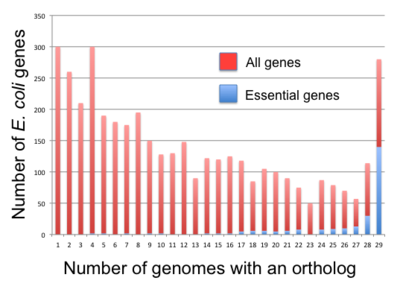
v bakterie, základní geny se zdají být konzervovanější než geny nepodstatné [71] ale korelace není příliš silná. Například pouze 34% z B. subtilis základní geny mají spolehlivost ortology celkově Firmicutes a 61% E-coli základní geny mají ve všech spolehlivé ortology Gama-proteobakterie.[70] Fang a kol. (2005) perzistentní geny jako geny přítomné ve více než 85% genomů kladu.[70] Našli 475 a 611 takových genů pro B. subtilis a E-coli, resp. Dále klasifikovali geny do pěti tříd podle perzistence a esenciality: perzistentní geny, esenciální geny, perzistentní neesenciální (PNE) geny (276 v B. subtilis409 palců E-coli), základní nepersperzní (ENP) geny (73 in.) B. subtilis, 33 palců E-coli) a netrvalé neesenciální (NPNE) geny (3558 palců) B. subtilis, 3 525 palců E-coli). Fang a kol. nalezeno 257 perzistentních genů, které existují jak v B. subtilis (pro Firmicutes) a E-coli (pro gama-proteobakterie). Z nich bylo 144 (respektive 139) dříve identifikováno jako nezbytné v B. subtilis (příslušně) E-coli) a 25 (respektive 18) z 257 genů není přítomno ve 475 B. subtilis (respektive 611 E-coli) perzistentní geny. Všichni ostatní členové skupiny jsou geny PNE.[70]
v eukaryoty, 83% ortologů jedna k jedné mezi Schizosaccharomyces pombe a Saccharomyces cerevisiae mají konzervovanou esenciálnost, to znamená, že jsou nepodstatné u obou druhů nebo esenciální u obou druhů. Zbývajících 17% genů je u jednoho druhu neesenciálních a u druhého esenciálních.[72] To je vzhledem k tomu pozoruhodné S. pombe je oddělena od S. cerevisiae přibližně 400 miliony let vývoje.[73]
Obecně platí, že vysoce konzervované a tedy starší geny (tj. Geny s dřívějším fylogenetickým původem) budou pravděpodobněji esenciální než mladší geny - i když byly duplikovány.[74]
Studie
Experimentální studium esenciálních genů je omezeno skutečností, že podle definice je inaktivace esenciálního genu pro organismus smrtelná. Proto je nelze jednoduše smazat nebo mutovat za účelem analýzy výsledku fenotypy (běžná technika v genetika ).
Existují však určité okolnosti, za kterých lze manipulovat s podstatnými geny. v diploidní organismy, může být zapotřebí pouze jedna funkční kopie některých základních genů (haplosufficiency ), přičemž heterozygot vykazoval poučný fenotyp. Některé základní geny mohou tolerovat mutace, které jsou škodlivé, ale nejsou zcela smrtelné, protože nezruší funkci genu úplně.
Výpočetní analýza může odhalit mnoho vlastností proteinů, aniž by je experimentálně analyzoval, např. při pohledu na homologní proteiny, funkce, struktura atd. (viz také níže, Předpovídání základních genů). Produkty základních genů lze také studovat, když exprimován v jiných organismech, nebo při čištění a studiu in vitro.
Podmíněně esenciální geny jsou snadněji studovatelné. Byly identifikovány teplotně citlivé varianty esenciálních genů, které kódují produkty, které ztrácejí funkci při vysokých teplotách, a tak vykazují fenotyp pouze při zvýšené teplotě.[75]
Reprodukovatelnost
Pokud se testy na základní geny opakují v nezávislých laboratořích, často vedou k různým seznamům genů. Například obrazovky v E-coli přinesly od ~ 300 do ~ 600 esenciálních genů (viz stůl 1). Tyto rozdíly jsou ještě výraznější, pokud se používají různé bakteriální kmeny (viz Obrázek 2). Běžným vysvětlením je, že experimentální podmínky jsou odlišné nebo že povaha mutace může být odlišná (např. Úplná delece genu vs. mutant transposonu).[4] Zejména transposonové obrazovky se těžko reprodukují, vzhledem k tomu, že transposon se může vložit do mnoha pozic v genu. Inzerce na 3 'konec esenciálního genu nemusí mít smrtelný fenotyp (nebo vůbec žádný fenotyp), a proto nemusí být jako takové rozpoznána. To může vést k chybným anotacím (zde: falešné negativy).[76]
Srovnání CRISPR / cas9 a RNAi obrazovky. Obrazovky k identifikaci základních genů u člověka chronická myelogenní leukémie buněčná linie K562 s těmito dvěma metodami vykazovala pouze omezené překrývání. Při 10% míře falešně pozitivních výsledků bylo na obrazovce Cas9 identifikováno ~ 4 500 genů oproti ~ 3 100 v shRNA obrazovce, v obou bylo identifikováno pouze ~ 1 200 genů.[77]
Různé esenciální geny v různých organismech
Různé organismy mohou mít různé esenciální geny. Například, Bacillus subtilis má 271 esenciálních genů.[20] Asi polovina (150) z ortologický geny v E-coli jsou také zásadní. Dalších 67 genů, které jsou nezbytné E-coli nejsou v B. subtilis, zatímco 86 E-coli základní geny nemají žádné B. subtilis ortolog.[24] v Mycoplasma genitalium nejméně 18 genů je esenciálních, u kterých není esenciálních M. bovis.[78] Mnoho z těchto různých základních genů je způsobeno paralogy nebo alternativními metabolickými cestami.[1]
Takové různé základní geny v bakteriích mohou být použity k vývoji cílené antibakteriální terapie proti určitým specifickým patogenům, aby se snížila odolnost proti antibiotikům v době mikrobiomu.[79] Stone a kol. (2015) použili rozdíl v základních genech v bakteriích k vývoji selektivních léků proti orálnímu patogenu Porphyromonas gingivalis, spíše než prospěšné bakterie Streptococcus sanguis.[80]
Předpověď
Esenciální geny lze předvídat výpočetně. Většina metod však do určité míry využívá experimentální data („tréninkové sady“). Chen a kol.[81] stanovila čtyři kritéria pro výběr tréninkových sad pro tyto předpovědi: (1) základní geny ve vybrané tréninkové sestavě by měly být spolehlivé; (2) podmínky růstu, ve kterých jsou definovány základní geny, by měly být v tréninkových a predikčních sadách konzistentní; (3) druhy používané jako výcviková sada by měly úzce souviset s cílovým organismem; a (4) organismy používané jako tréninkové a predikční sady by měly vykazovat podobné fenotypy nebo životní styl. Zjistili také, že velikost tréninkové sady by měla být alespoň 10% z celkových genů, aby se získaly přesné předpovědi. Některé přístupy k předpovědi základních genů jsou:
Srovnávací genomika. Krátce po prvních genomech (z Haemophilus influenzae a Mycoplasma genitalium ) byly k dispozici, Mushegian et al.[82] se pokusil předpovědět počet základních genů na základě běžných genů u těchto dvou druhů. Předpokládalo se, že na dlouhou evoluční vzdálenost, která odděluje tyto dvě bakterie, by měly být zachovány pouze základní geny. Tato studie identifikovala přibližně 250 kandidátních esenciálních genů.[82] Jak bylo k dispozici více genomů, počet předpokládaných esenciálních genů se stále zmenšoval, protože více genomů sdílelo méně a méně genů. V důsledku toho byl učiněn závěr, že univerzální konzervované jádro sestává z méně než 40 genů.[83][84] Tato sada konzervovaných genů však není totožná se sadou esenciálních genů, protože různé druhy se spoléhají na různé esenciální geny.
Podobný přístup byl použit k odvození základních genů z pan-genom z Brucella druh. 42 kompletní Brucella genomy a celkem 132 143 genů kódujících proteiny bylo použito k předpovědi 1252 potenciálních esenciálních genů, odvozených od jádra genomu ve srovnání s prokaryotickou databází esenciálních genů.[85]
Síťová analýza. Po první interakci proteinových sítí droždí byly zveřejněny,[86] bylo zjištěno, že vysoce spojené proteiny (např interakce protein-protein ) jsou pravděpodobněji zásadní.[87] Vysoce spojené proteiny však mohou být experimentálními artefakty a vysoká konektivita může spíše představovat pleiotropie místo podstaty.[88] Síťové metody byly nicméně vylepšeny přidáním dalších kritérií, a proto mají určitou hodnotu při předpovídání základních genů.[89]
Strojové učení. Hua a kol. použitý Strojové učení předpovídat základní geny u 25 bakteriálních druhů.[90]
Hurstův index. Liu a kol. (2015)[91] použil Hurstův exponent, charakteristický parametr popisující korelaci dalekého dosahu v DNA k předpovědi základních genů. V 31 z 33 bakteriálních genomů byly hladiny významnosti Hurstových exponentů esenciálních genů významně vyšší než pro odpovídající celou sadu genů, zatímco hladiny významnosti Hurstových exponentů neesenciálních genů zůstaly nezměněny nebo se zvýšily jen mírně.
Minimální genomy. Rovněž se předpokládalo, že z nich lze odvodit základní geny minimální genomy které údajně obsahují pouze základní geny. Problém je v tom, že nejmenší genomy patří parazitickým (nebo symbiontovým) druhům, které mohou přežít se sníženou sadou genů, protože získávají mnoho živin od svých hostitelů. Například jedním z nejmenších genomů je genom Hodgkinia cicadicola, a symbiont cikád, obsahující pouze 144 kB DNA kódující pouze 188 genů.[92] Stejně jako ostatní symbionty, Hodgkinia přijímá mnoho svých živin od svého hostitele, takže jeho geny nemusí být nezbytné.
Metabolické modelování. Esenciální geny lze také předpovědět v kompletně sekvenovaných genomech pomocí metabolická rekonstrukce, tj. rekonstrukcí úplného metabolismu z obsahu genů a následnou identifikací těch genů a cest, u kterých bylo u jiných druhů zjištěno, že jsou nezbytné. Tato metoda však může být narušena proteiny neznámé funkce. Mnoho organismů má navíc záložní nebo alternativní cesty, které je třeba vzít v úvahu (viz obrázek 1). Metabolické modelování použil také Basler (2015) k vývoji metody pro predikci základních metabolických genů.[93] Analýza rovnováhy toků, metoda metabolického modelování, byla nedávno použita k predikci esenciálních genů v metabolismu jasného buněčného karcinomu ledvinových buněk.[94]
Geny neznámé funkce. Překvapivě nemá významný počet esenciálních genů žádnou známou funkci. Například mezi 385 základními kandidáty v M. genitalium, žádná funkce nemohla být připsána 95 genům[6] i když se tento počet do roku 2011 snížil na 75.[84] Většina neznámých funkčně esenciálních genů má potenciální biologické funkce související s jednou ze tří základních funkcí.[1]
ZUPLS. Song a kol. představil novou metodu k předpovědi esenciálních genů, která využívá pouze křivku Z a další funkce založené na sekvenci.[95] Tyto rysy lze snadno vypočítat ze sekvencí DNA / aminokyselin. Spolehlivost této metody však zůstává trochu nejasná.
Servery pro predikci základních genů. Guo a kol. (2015) vyvinuli tři online služby pro predikci základních genů v bakteriálních genomech. Tyto volně dostupné nástroje jsou použitelné pro sekvence jednotlivých genů bez anotovaných funkcí, jednotlivé geny s určitými názvy a úplné genomy bakteriálních kmenů.[96] Kong a kol. (2019) vyvinuli ePath databáze, kterou lze použít k prohledání> 4 000 bakteriálních druhů pro predikci základních genů.[41]
Esenciální proteinové domény
Ačkoli většina esenciálních genů kóduje proteiny, mnoho esenciálních proteinů se skládá z jedné domény. Tato skutečnost byla použita k identifikaci esenciálních proteinových domén. Goodacre a kol. identifikovali stovky zásadních domény neznámé funkce (eDUF).[97] Lu a kol.[98] představil podobný přístup a identifikoval 3 450 domén, které jsou nezbytný alespoň u jednoho mikrobiálního druhu.
Viz také
Reference
- ^ A b C d E F G h i j k Xu, Ping; Ge, Xiuchun; Chen, Lei; Wang, Xiaojing; Dou, Yuetan; Xu, Jerry Z .; Patel, Jenishkumar R .; Stone, Victoria; Trinh, My; Evans, Karra; Kitten, Todd (prosinec 2011). „Identifikace esenciálního genu v celém genomu u Streptococcus sanguinis“. Vědecké zprávy. 1 (1): 125. Bibcode:2011NatSR ... 1E.125X. doi:10.1038 / srep00125. ISSN 2045-2322. PMC 3216606. PMID 22355642.
- ^ A b Zhang R, Lin Y (leden 2009). „DEG 5.0, databáze základních genů u prokaryot i eukaryot“. Výzkum nukleových kyselin. 37 (Problém s databází): D455-8. doi:10.1093 / nar / gkn858. PMC 2686491. PMID 18974178.
- ^ Sassetti CM, Boyd DH, Rubin EJ (2003). "Geny potřebné pro růst mykobakterií definované mutagenezí s vysokou hustotou". Mol Microbiol. 48 (1): 77–84. doi:10.1046 / j.1365-2958.2003.03425.x. PMID 12657046.
- ^ A b C d Gerdes S, Edwards R, Kubal M, Fonstein M, Stevens R, Osterman A (říjen 2006). "Základní geny na metabolických mapách". Aktuální názor na biotechnologie. 17 (5): 448–56. doi:10.1016 / j.copbio.2006.08.006. PMID 16978855.
- ^ Hutchison CA, Peterson SN, Gill SR, Cline RT, White O, Fraser CM, Smith HO, Venter JC (prosinec 1999). "Globální transpozonová mutageneze a minimální genom Mycoplasma". Věda. 286 (5447): 2165–9. doi:10.1126 / science.286.5447.2165. PMID 10591650. S2CID 235447.
- ^ A b Glass JI, Assad-Garcia N, Alperovich N, Yooseph S, Lewis MR, Maruf M, Hutchison CA, Smith HO, Venter JC (leden 2006). „Esenciální geny minimální bakterie“. Sborník Národní akademie věd Spojených států amerických. 103 (2): 425–30. Bibcode:2006PNAS..103..425G. doi:10.1073 / pnas.0510013103. PMC 1324956. PMID 16407165.
- ^ Ji Y, Zhang B, Van SF, Warren P, Woodnutt G, Burnham MK, Rosenberg M (září 2001). "Identifikace kritických stafylokokových genů pomocí podmíněných fenotypů generovaných antisense RNA". Věda. 293 (5538): 2266–9. Bibcode:2001Sci ... 293.2266J. doi:10.1126 / science.1063566. PMID 11567142. S2CID 24126939.
- ^ Forsyth RA, Haselbeck RJ, Ohlsen KL, Yamamoto RT, Xu H, Trawick JD, Wall D, Wang L, Brown-Driver V, Froelich JM, C KG, King P, McCarthy M, Malone C, Misiner B, Robbins D, Tan Z, Zhu Zy ZY, Carr G, Mosca DA, Zamudio C, Foulkes JG, Zyskind JW (březen 2002). „Strategie pro identifikaci základních genů v Staphylococcus aureus v rámci celého genomu.“ Molekulární mikrobiologie. 43 (6): 1387–400. doi:10.1046 / j.1365-2958.2002.02832.x. PMID 11952893.
- ^ Akerley BJ, Rubin EJ, Novick VL, Amaya K, Judson N, Mekalanos JJ (leden 2002). „Analýza v genomové škále pro identifikaci genů potřebných pro růst nebo přežití Haemophilus influenzae“. Sborník Národní akademie věd Spojených států amerických. 99 (2): 966–71. Bibcode:2002PNAS ... 99..966A. doi:10.1073 / pnas.012602299. PMC 117414. PMID 11805338.
- ^ Thanassi JA, Hartman-Neumann SL, Dougherty TJ, Dougherty BA, Pucci MJ (červenec 2002). „Identifikace 113 konzervovaných esenciálních genů pomocí vysoce výkonného systému narušení genů u Streptococcus pneumoniae“. Výzkum nukleových kyselin. 30 (14): 3152–62. doi:10.1093 / nar / gkf418. PMC 135739. PMID 12136097.
- ^ Song JH, Ko KS, Lee JY, Baek JY, Oh WS, Yoon HS, Jeong JY, Chun J (červen 2005). "Identifikace základních genů u Streptococcus pneumoniae alelickou substituční mutagenezí". Molekuly a buňky. 19 (3): 365–74. PMID 15995353.
- ^ A b Le Breton Y, Belew AT, Valdes KM, Islam E, Curry P, Tettelin H, Shirtliff ME, El-Sayed NM, McIver KS (květen 2015). „Základní geny v základním genomu lidského patogenu Streptococcus pyogenes“. Vědecké zprávy. 5: 9838. Bibcode:2015NatSR ... 5E9838L. doi:10.1038 / srep09838. PMC 4440532. PMID 25996237.
- ^ Chen L, Ge X, Xu P (2015). "Identifikace základních genů Streptococcus sanguinis pomocí mutace delece v celém genomu". Genová esence. Metody v molekulární biologii. 1279. str. 15–23. doi:10.1007/978-1-4939-2398-4_2. ISBN 978-1-4939-2397-7. PMC 4819415. PMID 25636610.
- ^ Sassetti CM, Boyd DH, Rubin EJ (říjen 2001). „Komplexní identifikace podmíněně esenciálních genů v mykobakteriích“. Sborník Národní akademie věd Spojených států amerických. 98 (22): 12712–7. Bibcode:2001PNAS ... 9812712S. doi:10.1073 / pnas.231275498. PMC 60119. PMID 11606763.
- ^ Lamichhane G, Freundlich JS, Ekins S, Wickramaratne N, Nolan ST, Bishai WR (únor 2011). „Základní metabolity Mycobacterium tuberculosis a jejich napodobeniny“. mBio. 2 (1): e00301-10. doi:10,1 128 / mBio.00301-10. PMC 3031304. PMID 21285434.
- ^ A b Griffin JE, Gawronski JD, Dejesus MA, Ioerger TR, Akerley BJ, Sassetti CM (září 2011). „Fenotypové profilování s vysokým rozlišením definuje geny nezbytné pro růst mykobakterií a katabolismus cholesterolu“. PLOS patogeny. 7 (9): e1002251. doi:10.1371 / journal.ppat.1002251. PMC 3182942. PMID 21980284.
- ^ Long JE, DeJesus M, Ward D, Baker RE, Ioerger T, Sassetti CM (2015). "Identifikace základních genů v Mycobacterium tuberculosis pomocí globálního fenotypového profilování". Genová esence. Metody v molekulární biologii. 1279. str. 79–95. doi:10.1007/978-1-4939-2398-4_6. ISBN 978-1-4939-2397-7. PMID 25636614.
- ^ DeJesus MA, Gerrick ER, Xu W, Park SW, Long JE, Boutte CC, Rubin EJ, Schnappinger D, Ehrt S, Fortune SM, Sassetti CM, Ioerger TR (leden 2017). „Komplexní analýza esenciálnosti genomu Mycobacterium tuberculosis nasycenou transpozonovou mutagenezí“. mBio. 8 (1): e02133–16. doi:10,1 128 / mBio.02133-16. PMC 5241402. PMID 28096490.
- ^ Ghosh S, Baloni P, Mukherjee S, Anand P, Chandra N (prosinec 2013). „Víceúrovňový víceúrovňový přístup ke studiu základních genů v Mycobacterium tuberculosis“. Biologie systémů BMC. 7: 132. doi:10.1186/1752-0509-7-132. PMC 4234997. PMID 24308365.
- ^ A b Kobayashi K, Ehrlich SD, Albertini A, Amati G, Andersen KK, Arnaud M a kol. (Duben 2003). „Esenciální geny Bacillus subtilis“. Sborník Národní akademie věd Spojených států amerických. 100 (8): 4678–83. Bibcode:2003PNAS..100,4678K. doi:10.1073 / pnas.0730515100. PMC 153615. PMID 12682299.
- ^ A b Commichau FM, Pietack N, Stülke J (červen 2013). „Esenciální geny v Bacillus subtilis: přehodnocení po deseti letech“. Molekulární biosystémy. 9 (6): 1068–75. doi:10.1039 / c3mb25595f. PMID 23420519. S2CID 23769853.
- ^ Gerdes SY, Scholle MD, Campbell JW, Balázsi G, Ravasz E, Daugherty MD a kol. (Říjen 2003). "Experimentální stanovení a analýza na úrovni systému základních genů v Escherichia coli MG1655". Journal of Bacteriology. 185 (19): 5673–84. doi:10.1128 / JB.185.19.5673-5684.2003. PMC 193955. PMID 13129938.
- ^ Kang Y, Durfee T, Glasner JD, Qiu Y, Frisch D, Winterberg KM a kol. (Srpen 2004). "Systematická mutageneze genomu Escherichia coli". Journal of Bacteriology. 186 (15): 4921–30. doi:10.1128 / JB.186.15.4921-4930.2004. PMC 451658. PMID 15262929.
- ^ A b Baba T, Ara T, Hasegawa M, Takai Y, Okumura Y, Baba M a kol. (2006). „Konstrukce Escherichia coli K-12 in-frame, single-gen knockout mutantů: kolekce Keio“. Molekulární biologie systémů. 2: 2006.0008. doi:10.1038 / msb4100050. PMC 1681482. PMID 16738554.
- ^ Jacobs MA, Alwood A, Thaipisuttikul I, Spencer D, Haugen E, Ernst S a kol. (Listopad 2003). „Komplexní knihovna mutantů transpozonů Pseudomonas aeruginosa“. Sborník Národní akademie věd Spojených států amerických. 100 (24): 14339–44. Bibcode:2003PNAS..10014339J. doi:10.1073 / pnas.2036282100. PMC 283593. PMID 14617778.
- ^ Hutcherson JA, Gogeneni H, Yoder-Himes D, Hendrickson EL, Hackett M, Whiteley M a kol. (Srpen 2016). „Srovnání inherentně esenciálních genů Porphyromonas gingivalis identifikovaných ve dvou knihovnách sekvenujících transpozony“. Molekulární orální mikrobiologie. 31 (4): 354–64. doi:10.1111 / omi.12135. PMC 4788587. PMID 26358096.
- ^ Liberati NT, Urbach JM, Miyata S, Lee DG, Drenkard E, Wu G a kol. (Únor 2006). „Objednaná neredundantní knihovna mutantů inzerujících transpozon PA14 kmene Pseudomonas aeruginosa“. Sborník Národní akademie věd Spojených států amerických. 103 (8): 2833–8. Bibcode:2006PNAS..103,2833L. doi:10.1073 / pnas.0511100103. PMC 1413827. PMID 16477005.
- ^ Knuth K, Niesalla H, Hueck CJ, Fuchs TM (březen 2004). "Rozsáhlá identifikace základních genů Salmonella zachycením letálních inzercí". Molekulární mikrobiologie. 51 (6): 1729–44. doi:10.1046 / j.1365-2958.2003.03944.x. PMID 15009898.
- ^ Salama NR, Shepherd B, Falkow S (prosinec 2004). „Globální transpozonová mutageneze a analýza základních genů Helicobacter pylori“. Journal of Bacteriology. 186 (23): 7926–35. doi:10.1128 / JB.186.23.7926-7935.2004. PMC 529078. PMID 15547264.
- ^ Suzuki N, Inui M, Yukawa H (2011). Vysoce výkonná transpozonová mutageneze Corynebacterium glutamicum. Metody v molekulární biologii. 765. 409–17. doi:10.1007/978-1-61779-197-0_24. ISBN 978-1-61779-196-3. PMID 21815106.
- ^ Gallagher LA, Ramage E, Jacobs MA, Kaul R, Brittnacher M, Manoil C (leden 2007). „Komplexní knihovna mutantů transpozonů Francisella novicida, zástupce biologických zbraní“. Sborník Národní akademie věd Spojených států amerických. 104 (3): 1009–14. Bibcode:2007PNAS..104,1009G. doi:10.1073 / pnas.0606713104. PMC 1783355. PMID 17215359.
- ^ Stahl M, Stintzi A (červen 2011). „Identifikace esenciálních genů v genomu C. jejuni zdůrazňuje oblasti hyper-variabilní plasticity“. Funkční a integrační genomika. 11 (2): 241–57. doi:10.1007 / s10142-011-0214-7. PMID 21344305. S2CID 24054117.
- ^ Stahl M, Stintzi A (2015). "Microarray sledování transpozonu pro mapování podmíněně esenciálních genů v Campylobacter jejuni". Genová esence. Metody v molekulární biologii. 1279. s. 1–14. doi:10.1007/978-1-4939-2398-4_1. ISBN 978-1-4939-2397-7. PMID 25636609.
- ^ French CT, Lao P, Loraine AE, Matthews BT, Yu H, Dybvig K (červenec 2008). „Velká transpozonová mutageneze Mycoplasma pulmonis“. Molekulární mikrobiologie. 69 (1): 67–76. doi:10.1111 / j.1365-2958.2008.06262.x. PMC 2453687. PMID 18452587.
- ^ Cameron DE, Urbach JM, Mekalanos JJ (červen 2008). „Definovaná knihovna transpozonových mutantů a její použití při identifikaci genů motility u Vibrio cholerae“. Sborník Národní akademie věd Spojených států amerických. 105 (25): 8736–41. Bibcode:2008PNAS..105,8736C. doi:10.1073 / pnas.0803281105. PMC 2438431. PMID 18574146.
- ^ Langridge GC, Phan MD, Turner DJ, Perkins TT, Parts L, Haase J a kol. (Prosinec 2009). "Simultánní analýza každého genu Salmonella Typhi s použitím jednoho milionu transposonových mutantů". Výzkum genomu. 19 (12): 2308–16. doi:10.1101 / gr.097097.109. PMC 2792183. PMID 19826075.
- ^ Chaudhuri RR, Allen AG, Owen PJ, Shalom G, Stone K, Harrison M a kol. (Červenec 2009). „Komplexní identifikace základních genů Staphylococcus aureus pomocí Transposonem zprostředkované diferenciální hybridizace (TMDH)“. BMC Genomics. 10: 291. doi:10.1186/1471-2164-10-291. PMC 2721850. PMID 19570206.
- ^ Christen B, Abeliuk E, Collier JM, Kalogeraki VS, Passarelli B, Coller JA, Fero MJ, McAdams HH, Shapiro L (srpen 2011). „Základní genom bakterie“. Molekulární biologie systémů. 7: 528. doi:10.1038 / msb.2011.58. PMC 3202797. PMID 21878915.
- ^ Mendum TA, Newcombe J, Mannan AA, Kierzek AM, McFadden J (prosinec 2011). „Výslech údajů o globální mutagenezi pomocí modelu genomu Neisseria meningitidis v měřítku genomu k posouzení genové kondice in vitro a v séru“. Genome Biology. 12 (12): R127. doi:10.1186/gb-2011-12-12-r127. PMC 3334622. PMID 22208880.
- ^ Kuehl JV, Price MN, Ray J, Wetmore KM, Esquivel Z, Kazakov AE, et al. (Květen 2014). "Functional genomics with a comprehensive library of transposon mutants for the sulfate-reducing bacterium Desulfovibrio alaskensis G20". mBio. 5 (3): e01041-14. doi:10.1128/mBio.01041-14. PMC 4045070. PMID 24865553.
- ^ A b Kong, Xiangzhen; Zhu, Bin; Stone, Victoria N.; Ge, Xiuchun; El-Rami, Fadi E.; Donghai, Huangfu; Xu, Ping (December 2019). "ePath: an online database towards comprehensive essential gene annotation for prokaryotes". Vědecké zprávy. 9 (1): 12949. Bibcode:2019NatSR...912949K. doi:10.1038/s41598-019-49098-w. ISSN 2045-2322. PMC 6737131. PMID 31506471.
- ^ A b Kim DU, Hayles J, Kim D, Wood V, Park HO, Won M, et al. (Červen 2010). "Analysis of a genome-wide set of gene deletions in the fission yeast Schizosaccharomyces pombe". Přírodní biotechnologie. 28 (6): 617–623. doi:10.1038/nbt.1628. PMC 3962850. PMID 20473289.
- ^ A b Kamath RS, Fraser AG, Dong Y, Poulin G, Durbin R, Gotta M, et al. (Leden 2003). "Systematic functional analysis of the Caenorhabditis elegans genome using RNAi". Příroda. 421 (6920): 231–7. Bibcode:2003Natur.421..231K. doi:10.1038/nature01278. hdl:10261/63159. PMID 12529635. S2CID 15745225.
- ^ A b Spradling AC, Stern D, Beaton A, Rhem EJ, Laverty T, Mozden N, et al. (Září 1999). "The Berkeley Drosophila Genome Project gene disruption project: Single P-element insertions mutating 25% of vital Drosophila genes". Genetika. 153 (1): 135–77. PMC 1460730. PMID 10471706.
- ^ A b Amsterdam A, Nissen RM, Sun Z, Swindell EC, Farrington S, Hopkins N (August 2004). "Identification of 315 genes essential for early zebrafish development". Sborník Národní akademie věd Spojených států amerických. 101 (35): 12792–7. Bibcode:2004PNAS..10112792A. doi:10.1073/pnas.0403929101. PMC 516474. PMID 15256591.
- ^ White JK, Gerdin AK, Karp NA, Ryder E, Buljan M, Bussell JN, et al. (Červenec 2013). „Generace v celém genomu a systematické fenotypování knockoutovaných myší odhaluje nové role mnoha genů“. Buňka. 154 (2): 452–64. doi:10.1016 / j.cell.2013.06.022. PMC 3717207. PMID 23870131.
- ^ A b Liao BY, Zhang J (May 2008). "Null mutations in human and mouse orthologs frequently result in different phenotypes". Sborník Národní akademie věd Spojených států amerických. 105 (19): 6987–92. Bibcode:2008PNAS..105.6987L. doi:10.1073/pnas.0800387105. PMC 2383943. PMID 18458337.
- ^ Georgi B, Voight BF, Bućan M (květen 2013). Flint J (ed.). „Od myší k člověku: evoluční genomická analýza lidských ortologů základních genů“. Genetika PLOS. 9 (5): e1003484. doi:10.1371 / journal.pgen.1003484. PMC 3649967. PMID 23675308.
- ^ A b Sulem P, Helgason H, Oddson A, Stefansson H, Gudjonsson SA, Zink F, et al. (Květen 2015). "Identification of a large set of rare complete human knockouts". Genetika přírody. 47 (5): 448–52. doi:10.1038/ng.3243. PMID 25807282. S2CID 205349719.
- ^ A b Narasimhan VM, Hunt KA, Mason D, Baker CL, Karczewski KJ, Barnes MR, et al. (Duben 2016). "Health and population effects of rare gene knockouts in adult humans with related parents". Věda. 352 (6284): 474–7. Bibcode:2016Sci...352..474N. doi:10.1126/science.aac8624. PMC 4985238. PMID 26940866.
- ^ Luo H, Lin Y, Gao F, Zhang CT, Zhang R (January 2014). "DEG 10, an update of the database of essential genes that includes both protein-coding genes and noncoding genomic elements". Výzkum nukleových kyselin. 42 (Database issue): D574-80. doi:10.1093/nar/gkt1131. PMC 3965060. PMID 24243843.
- ^ Tzafrir I, Pena-Muralla R, Dickerman A, Berg M, Rogers R, Hutchens S, et al. (Červenec 2004). "Identification of genes required for embryo development in Arabidopsis". Fyziologie rostlin. 135 (3): 1206–20. doi:10.1104/pp.104.045179. PMC 519041. PMID 15266054.
- ^ Wang T, Birsoy K, Hughes NW, Krupczak KM, Post Y, Wei JJ, et al. (Listopad 2015). "Identification and characterization of essential genes in the human genome". Věda. 350 (6264): 1096–101. Bibcode:2015Sci...350.1096W. doi:10.1126/science.aac7041. PMC 4662922. PMID 26472758.
- ^ Blomen VA, Májek P, Jae LT, Bigenzahn JW, Nieuwenhuis J, Staring J, et al. (Listopad 2015). "Gene essentiality and synthetic lethality in haploid human cells". Věda. 350 (6264): 1092–6. Bibcode:2015Sci...350.1092B. doi:10.1126/science.aac7557. PMID 26472760. S2CID 26529733.
- ^ Georgi B, Voight BF, Bućan M (květen 2013). „Od myší k člověku: evoluční genomická analýza lidských ortologů základních genů“. Genetika PLOS. 9 (5): e1003484. doi:10.1371 / journal.pgen.1003484. PMC 3649967. PMID 23675308.
- ^ Liao BY, Zhang J (August 2007). "Mouse duplicate genes are as essential as singletons". Trendy v genetice. 23 (8): 378–81. doi:10.1016/j.tig.2007.05.006. PMID 17559966.
- ^ Mewes HW, Frishman D, Güldener U, Mannhaupt G, Mayer K, Mokrejs M, et al. (Leden 2002). "MIPS: a database for genomes and protein sequences". Výzkum nukleových kyselin. 30 (1): 31–4. doi:10.1093/nar/30.1.31. PMC 99165. PMID 11752246.
- ^ A b Giaever G, Chu AM, Ni L, Connelly C, Riles L, Véronneau S, et al. (Červenec 2002). "Functional profiling of the Saccharomyces cerevisiae genome". Příroda. 418 (6896): 387–91. Bibcode:2002Natur.418..387G. doi:10.1038/nature00935. PMID 12140549. S2CID 4400400.
- ^ Yu D, Silva MC, Shenk T (October 2003). "Functional map of human cytomegalovirus AD169 defined by global mutational analysis". Sborník Národní akademie věd Spojených států amerických. 100 (21): 12396–401. Bibcode:2003PNAS..10012396Y. doi:10.1073/pnas.1635160100. PMC 218769. PMID 14519856.
- ^ A b Dobson BM, Tscharke DC (November 2015). "Redundancy complicates the definition of essential genes for vaccinia virus". The Journal of General Virology. 96 (11): 3326–37. doi:10.1099/jgv.0.000266. PMC 5972330. PMID 26290187.
- ^ Dedrick RM, Marinelli LJ, Newton GL, Pogliano K, Pogliano J, Hatfull GF (May 2013). "Functional requirements for bacteriophage growth: gene essentiality and expression in mycobacteriophage Giles". Molekulární mikrobiologie. 88 (3): 577–89. doi:10,1111 / mmi.12210. PMC 3641587. PMID 23560716.
- ^ Thomas JA, Benítez Quintana AD, Bosch MA, Coll De Peña A, Aguilera E, Coulibaly A, et al. (Listopad 2016). "Identification of Essential Genes in the Salmonella Phage SPN3US Reveals Novel Insights into Giant Phage Head Structure and Assembly". Journal of Virology. 90 (22): 10284–10298. doi:10.1128/JVI.01492-16. PMC 5105663. PMID 27605673.
- ^ Pál C, Papp B, Lercher MJ, Csermely P, Oliver SG, Hurst LD (March 2006). "Chance and necessity in the evolution of minimal metabolic networks". Příroda. 440 (7084): 667–70. Bibcode:2006Natur.440..667P. doi:10.1038/nature04568. PMID 16572170. S2CID 4424895.
- ^ Mori H, Baba T, Yokoyama K, Takeuchi R, Nomura W, Makishi K, Otsuka Y, Dose H, Wanner BL (2015). "Identification of essential genes and synthetic lethal gene combinations in Escherichia coli K-12". Gene Essentiality. Metody v molekulární biologii. 1279. str. 45–65. doi:10.1007/978-1-4939-2398-4_4. ISBN 978-1-4939-2397-7. PMID 25636612.
- ^ Verhagen LM, de Jonge MI, Burghout P, Schraa K, Spagnuolo L, Mennens S, Eleveld MJ, van der Gaast-de Jongh CE, Zomer A, Hermans PW, Bootsma HJ (2014). "Genome-wide identification of genes essential for the survival of Streptococcus pneumoniae in human saliva". PLOS ONE. 9 (2): e89541. Bibcode:2014PLoSO...989541V. doi:10.1371/journal.pone.0089541. PMC 3934895. PMID 24586856.
- ^ D'Souza G, Kost C (November 2016). "Experimental Evolution of Metabolic Dependency in Bacteria". Genetika PLOS. 12 (11): e1006364. doi:10.1371/journal.pgen.1006364. PMC 5096674. PMID 27814362.
- ^ Xu, Ping; Widmer, Giovanni; Wang, Yingping; Ozaki, Luiz S.; Alves, Joao M.; Serrano, Myrna G.; Puiu, Daniela; Manque, Patricio; Akiyoshi, Donna; Mackey, Aaron J.; Pearson, William R. (October 2004). "The genome of Cryptosporidium hominis". Příroda. 431 (7012): 1107–1112. Bibcode:2004Natur.431.1107X. doi:10.1038/nature02977. ISSN 0028-0836. PMID 15510150. S2CID 4394344.
- ^ Tipples G, McClarty G (June 1993). "The obligate intracellular bacterium Chlamydia trachomatis is auxotrophic for three of the four ribonucleoside triphosphates". Molekulární mikrobiologie. 8 (6): 1105–14. doi:10.1111/j.1365-2958.1993.tb01655.x. PMID 8361355.
- ^ Woods S, Coghlan A, Rivers D, Warnecke T, Jeffries SJ, Kwon T, et al. (Květen 2013). Sternberg PW (ed.). "Duplication and retention biases of essential and non-essential genes revealed by systematic knockdown analyses". Genetika PLOS. 9 (5): e1003330. doi:10.1371/journal.pgen.1003330. PMC 3649981. PMID 23675306.
- ^ A b C d Fang G, Rocha E, Danchin A (November 2005). "How essential are nonessential genes?". Molekulární biologie a evoluce. 22 (11): 2147–56. doi:10.1093/molbev/msi211. PMID 16014871.
- ^ Jordan IK, Rogozin IB, Wolf YI, Koonin EV (June 2002). "Essential genes are more evolutionarily conserved than are nonessential genes in bacteria". Výzkum genomu. 12 (6): 962–8. doi:10.1101/gr.87702. PMC 1383730. PMID 12045149.
- ^ Ryan CJ, Krogan NJ, Cunningham P, Cagney G (2013). "All or nothing: protein complexes flip essentiality between distantly related eukaryotes". Biologie genomu a evoluce. 5 (6): 1049–59. doi:10.1093 / gbe / evt074. PMC 3698920. PMID 23661563.
- ^ Sipiczki M (2000). "Where does fission yeast sit on the tree of life?". Genome Biology. 1 (2): REVIEWS1011. doi:10.1186/gb-2000-1-2-reviews1011. PMC 138848. PMID 11178233.
- ^ Chen WH, Trachana K, Lercher MJ, Bork P (July 2012). "Younger genes are less likely to be essential than older genes, and duplicates are less likely to be essential than singletons of the same age". Molekulární biologie a evoluce. 29 (7): 1703–6. doi:10.1093/molbev/mss014. PMC 3375470. PMID 22319151.
- ^ Kofoed M, Milbury KL, Chiang JH, Sinha S, Ben-Aroya S, Giaever G, et al. (Červenec 2015). "An Updated Collection of Sequence Barcoded Temperature-Sensitive Alleles of Yeast Essential Genes". G3. 5 (9): 1879–87. doi:10.1534/g3.115.019174. PMC 4555224. PMID 26175450.
- ^ Deng J, Su S, Lin X, Hassett DJ, Lu LJ (2013). Kim PM (ed.). "A statistical framework for improving genomic annotations of prokaryotic essential genes". PLOS ONE. 8 (3): e58178. Bibcode:2013PLoSO...858178D. doi:10.1371/journal.pone.0058178. PMC 3592911. PMID 23520492.
- ^ Morgens DW, Deans RM, Li A, Bassik MC (June 2016). "Systematic comparison of CRISPR/Cas9 and RNAi screens for essential genes". Přírodní biotechnologie. 34 (6): 634–6. doi:10.1038/nbt.3567. PMC 4900911. PMID 27159373.
- ^ Sharma S, Markham PF, Browning GF (2014). "Genes found essential in other mycoplasmas are dispensable in Mycoplasma bovis". PLOS ONE. 9 (6): e97100. Bibcode:2014PLoSO...997100S. doi:10.1371/journal.pone.0097100. PMC 4045577. PMID 24897538.
- ^ Stone VN, Xu P (December 2017). "Targeted antimicrobial therapy in the microbiome era". Molekulární orální mikrobiologie. 32 (6): 446–454. doi:10.1111/omi.12190. PMC 5697594. PMID 28609586.
- ^ Stone VN, Parikh HI, El-rami F, Ge X, Chen W, Zhang Y, et al. (06.11.2015). Merritt J (ed.). "Identification of Small-Molecule Inhibitors against Meso-2, 6-Diaminopimelate Dehydrogenase from Porphyromonas gingivalis". PLOS ONE. 10 (11): e0141126. Bibcode:2015PLoSO..1041126S. doi:10.1371/journal.pone.0141126. PMC 4636305. PMID 26544875.
- ^ Cheng J, Xu Z, Wu W, Zhao L, Li X, Liu Y, Tao S (2014). "Training set selection for the prediction of essential genes". PLOS ONE. 9 (1): e86805. Bibcode:2014PLoSO...986805C. doi:10.1371/journal.pone.0086805. PMC 3899339. PMID 24466248.
- ^ A b Mushegian AR, Koonin EV (September 1996). "A minimal gene set for cellular life derived by comparison of complete bacterial genomes". Sborník Národní akademie věd Spojených států amerických. 93 (19): 10268–73. Bibcode:1996PNAS...9310268M. doi:10.1073/pnas.93.19.10268. PMC 38373. PMID 8816789.
- ^ Charlebois RL, Doolittle WF (December 2004). "Computing prokaryotic gene ubiquity: rescuing the core from extinction". Výzkum genomu. 14 (12): 2469–77. doi:10.1101/gr.3024704. PMC 534671. PMID 15574825.
- ^ A b Juhas M, Eberl L, Glass JI (October 2011). "Essence of life: essential genes of minimal genomes". Trendy v buněčné biologii. 21 (10): 562–8. doi:10.1016/j.tcb.2011.07.005. PMID 21889892.
- ^ Yang X, Li Y, Zang J, Li Y, Bie P, Lu Y, Wu Q (April 2016). "Analysis of pan-genome to identify the core genes and essential genes of Brucella spp". Molekulární genetika a genomika. 291 (2): 905–12. doi:10.1007/s00438-015-1154-z. PMID 26724943. S2CID 14565579.
- ^ Schwikowski B, Uetz P, Fields S (December 2000). "A network of protein-protein interactions in yeast". Přírodní biotechnologie. 18 (12): 1257–61. doi:10.1038/82360. PMID 11101803. S2CID 3009359.
- ^ Jeong H, Mason SP, Barabási AL, Oltvai ZN (May 2001). "Letalita a centralita v proteinových sítích". Příroda. 411 (6833): 41–2. arXiv:cond-mat / 0105306. Bibcode:2001 Natur.411 ... 41J. doi:10.1038/35075138. PMID 11333967. S2CID 258942.
- ^ Yu H, Braun P, Yildirim MA, Lemmens I, Venkatesan K, Sahalie J, et al. (Říjen 2008). „Vysoce kvalitní mapa interakce binárních proteinů v síti kvasinek. Věda. 322 (5898): 104–10. Bibcode:2008Sci ... 322..104Y. doi:10.1126 / science.1158684. PMC 2746753. PMID 18719252.
- ^ Li X, Li W, Zeng M, Zheng R, Li M (February 2019). "Network-based methods for predicting essential genes or proteins: a survey". Briefings in Bioinformatics. 21 (2): 566–583. doi:10.1093/bib/bbz017. PMID 30776072.
- ^ Hua HL, Zhang FZ, Labena AA, Dong C, Jin YT, Guo FB (2016-01-01). "An Approach for Predicting Essential Genes Using Multiple Homology Mapping and Machine Learning Algorithms". BioMed Research International. 2016: 7639397. doi:10.1155/2016/7639397. PMC 5021884. PMID 27660763.
- ^ Liu X, Wang B, Xu L (2015). "Statistical Analysis of Hurst Exponents of Essential/Nonessential Genes in 33 Bacterial Genomes". PLOS ONE. 10 (6): e0129716. Bibcode:2015PLoSO..1029716L. doi:10.1371/journal.pone.0129716. PMC 4466317. PMID 26067107.
- ^ McCutcheon JP, McDonald BR, Moran NA (July 2009). Matic I (ed.). "Origin of an alternative genetic code in the extremely small and GC-rich genome of a bacterial symbiont". Genetika PLOS. 5 (7): e1000565. doi:10.1371/journal.pgen.1000565. PMC 2704378. PMID 19609354.
- ^ Basler G (2015). "Computational prediction of essential metabolic genes using constraint-based approaches". Gene Essentiality. Metody v molekulární biologii. 1279. 183–204. doi:10.1007/978-1-4939-2398-4_12. ISBN 978-1-4939-2397-7. PMID 25636620.
- ^ Gatto F, Miess H, Schulze A, Nielsen J (June 2015). "Flux balance analysis predicts essential genes in clear cell renal cell carcinoma metabolism". Vědecké zprávy. 5: 10738. Bibcode:2015NatSR...5E0738G. doi:10.1038/srep10738. PMC 4603759. PMID 26040780.
- ^ Song K, Tong T, Wu F (April 2014). "Predicting essential genes in prokaryotic genomes using a linear method: ZUPLS". Integrativní biologie. 6 (4): 460–9. doi:10.1039/c3ib40241j. PMID 24603751.
- ^ Guo FB, Ye YN, Ning LW, Wei W (2015). "Three computational tools for predicting bacterial essential genes". Gene Essentiality. Metody v molekulární biologii. 1279. pp. 205–17. doi:10.1007/978-1-4939-2398-4_13. ISBN 978-1-4939-2397-7. PMID 25636621.
- ^ Goodacre NF, Gerloff DL, Uetz P (December 2013). "Protein domains of unknown function are essential in bacteria". mBio. 5 (1): e00744-13. doi:10.1128/mBio.00744-13. PMC 3884060. PMID 24381303.
- ^ Lu Y, Lu Y, Deng J, Lu H, Lu LJ (2015). "Discovering essential domains in essential genes". Gene Essentiality. Metody v molekulární biologii. 1279. pp. 235–45. doi:10.1007/978-1-4939-2398-4_15. ISBN 978-1-4939-2397-7. PMID 25636623.
Další čtení
- Gao F, Luo H, Zhang CT, Zhang R (2015). "Gene essentiality analysis based on DEG 10, an updated database of essential genes". Gene Essentiality. Metody v molekulární biologii. 1279. pp. 219–33. doi:10.1007/978-1-4939-2398-4_14. ISBN 978-1-4939-2397-7. PMID 25636622.
- Long JL, ed. (2015). Gene Essentiality - Springer Methods and Protocols. Metody v molekulární biologii. 1279. Humana Press. p. 248. doi:10.1007/978-1-4939-2398-4. ISBN 978-1-4939-2397-7. S2CID 27547825.