Ornitholestes - Ornitholestes
| Ornitholestes | |
|---|---|
 | |
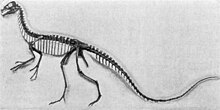
| Montáž na kostru, Královské muzeum Tyrrell | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Clade: | Dinosaurie |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Maniraptoromorpha |
| Podčeleď: | †Ornitholestinae Pavel, 1988 |
| Rod: | †Ornitholestes Osborn, 1903 |
| Zadejte druh | |
| †Ornitholestes hermanni Osborn, 1903 | |
Ornitholestes (což znamená „lupič ptáků“) je malý teropod dinosaurus pozdní jurský (Brushy Basin Člen z Morrison formace, prostřední Kimmeridgian věku, asi před 154 miliony let[1]) západní Laurasia (oblast, která se měla stát Severní Amerika ).
K datu, Ornitholestes je znám pouze z jedné částečné kostry se špatně rozdrcenou lebkou nalezenou u Lom na kabinu u Medicine Bow, Wyoming, v roce 1900. Popsal to Henry Fairfield Osborn v roce 1903. Neúplná ruka byla později připsána Ornitholestes, ačkoli se nyní zdá, že patří Tanycolagreus. The typ (a pouze známý) druh je O. hermanni. The konkrétní název ctí Americké muzeum přírodní historie preparátor Adam Hermann.
Popis

Ornitholestes byl bipedální masožravec.[2] Jeho hlava byla proporcionálně menší než u většiny ostatních dravých dinosaurů, ale lebka byla silně stavěná, s krátkým čenichem a robustní dolní čelistí.[3] The oběžné dráhy (oční důlky) byly poměrně velké a měřily přes 25 procent délky lebky.[4] Neexistuje žádný náznak a kostnatý oční kroužek.[5]
Přední zuby Ornitholestes byly poněkud kuželovité se sníženým ozubením; zadní zuby byly zakřivené a ostřeji zoubkované, podobně jako u jiných dinosaurů z teropodů.[6] Henry Fairfield Osborn (1903) počítal čtyři zuby premaxilla, jehož přední zub byl největší v horní čelisti.[7] V porovnání, Gregory S. Paul (1988) zobrazili lebku se zbývajícími třemi premaxilárními zuby, mnohem menší než ty, které ilustroval Osborn.[8] Každý horní čelist (hlavní nosná kost v horní čelisti) obsahovala deset zubů a každý zubař (kosti nesoucí zuby v dolní čelisti) obsahovaly dvanáct zubů.[9] Řady zubů Ornitholestes byly krátké, přičemž zubařská (spodní) řada byla ještě kratší než čelistní (horní) řada,[10] i když samotná zubní kost byla vzadu výjimečně dlouhá a dosahovala bodu pod středem oční důlky.[11] Zuby nepřesahovaly tak daleko dozadu jako oběžné dráhy a ani jedna řada zubů nepřesahovala mnohem více než jednu třetinu lebky.[12]

Zdá se, že oblast zlomené kosti poblíž vnějšího narisu (nosní dírky) vyklenuje nahoru, což vedlo Gregoryho S. Draví dinosauři světa (1988) Ornitholestes měl nosní roh "spíše jako kuřecí hřeben ve vzhledu."[13] Oba Oliver W.M. Rauhut (2003) a Kenneth Carpenter et al. (2005A) odmítl tuto interpretaci a naznačil, že vzplanutí kosti vzhůru bylo způsobeno postmortálním rozdrcením lebky.[14] Paulův aktualizovaný obrázek Ornitholestes v jeho 2010 Princeton Field Guide to Dinosaurs již neobsahuje nosní roh.[15]
Ornitholestes měl relativně krátký krk s mírnou sigmoidální křivkou (ve tvaru písmene S).[16] Ocas byl dlouhý a bičovitý, tvořil více než polovinu délky těla.[17] Ne všechny obratle byly zachovány, ale Osborn (1917) to odhadl Ornitholestes měl devět nebo deset krčních (krčních) obratlů, třináct hřbetních (zadních) obratlů, čtyři křížové (kyčle) obratle a 39 až 44 kaudálních (ocasních) obratlů.[18] Tesař et al. (2005A) zaznamenal, že vzorek obsahoval pět křížových obratlů.[19] Ornitholestes byl krátkonohý teropod, a to se odráželo v krátkých předozadních rozměrech krčních a hřbetních obratlů.[20]

Přední končetiny Ornitholestes byly relativně dlouhé, mírně pod dvě třetiny délky zadních nohou.[21] The humerus (horní část paže) byla silně stavěná a poněkud delší než poloměr a ulna (kosti předloktí).[22] Humerus a radius byly rovné.[23] Drápy na číslicích I a II ruky byly přibližně stejně velké.[24] Ačkoli se třetí nezměněný muž (drápová kost) nezachoval, extrapolace od nejbližších příbuzných Ornitholestes naznačuje, že to bylo pravděpodobně kratší než u prvních dvou.[24]
Ornitholestes je často zobrazován jako rychlý, dlouhonohý teropod, ale jeho kosti dolní končetiny byly poměrně krátké.[25] Osborn (1917) vypočítal, že holenní kost (holenní kost) byla pouze asi 70,6% tak dlouho, jak stehenní kost (stehenní kost).[26] The metatarzály (kosti nohy) byly umístěny těsně vedle sebe, ale nebyly spojeny dohromady.[27] Jak je u teropodů typické, chodidla byla tridaktylová (se třemi drápy s nosnými prsty).[28] John H. Ostrom (1969) poznamenal, že dráp číslice II (nejvnitřnější špička) byl větší než dráp číslic III a IV, a navrhl, že tato číslice mohla nést upravený srpový dráp podobný tomu z Deinonychus.[29] Jak však poznamenali Ostrom (1969) a Paul (1988), špatná ochrana číslice II ztěžuje potvrzení této hypotézy.[30]

Ve svém popisu z roku 1903 Osborn napsal, že délka Ornitholestes podél „lebky a páteře po obnovení“ bylo 2,22 m (7,28 ft).[31] Tato rekonstrukce však byla nepřesná a byla částečně založena na Othniel Charles Marsh obnova bazální sauropodomorf Anchisaurus a krk a trup byly příliš protáhlé.[32] David Norman (1985) a John Foster (2007) to odhadli Ornitholestes byl dlouhý asi 2 m (6,6 ft).[33] Gregory S. Paul 1988 Draví dinosauři světa uvedena délka Ornitholestes jako přibližně 2,08 m (6,82 ft).[34]
Paul (1988) a Foster (2007) to odhadují Ornitholestes vážil 12,6 kg (27,8 lb).[35] John A. Long a Peter Schouten (2008) navrhli mírně vyšší číslo, 15 kg (33 lb).[36]
Objev a pojmenování

Ornitholestes byl prvním teropodem, který byl objeven ve 20. letech 20. století.[37] The holotyp kostra (AMNH 619) byla vykopána v červenci 1900 v Lom na kabinu v Wyoming podle Americké muzeum přírodní historie expedice Petera C. Kaisena, Paula Millera a Frederic Brewster Loomis.[31][38] Představuje částečnou kostru s lebkou, včetně mnoha prvků páteře, předních končetin, pánve a zadních končetin. Henry Fairfield Osborn pojmenoval a vědecky popsal vzorek v roce 1903.[39] The rod název Ornitholestes, původně navrhl Theodore Gill, znamená „lupič ptáků“ a je odvozen z řecký ὄρνις/ornis, ornithos („pták“) a λῃστήσ/lestes ("lupič").[40] The druh název (O. hermanni) vyznamenává Adama Hermanna, hlavního preparátora v muzeu, který řídil restaurování a montáž kostry.[31]
Byla přidělena neúplná ruka (AMNH 587) Ornitholestes Osborn ve svém popisu rodu z roku 1903.[41] Jak však poznamenal Gregory S. Paul (1988), špatná ochrana odpovídajících prvků ve vzorku typu způsobila, že tato asociace byla „předběžná“.[42] V roce 2005 Kenneth Carpenter et al. popsal nový malý theropod, Tanycolagreus, jehož kostra byla nalezena v lomu Bone Cabin Quarry jen několik set metrů od AMNH 587.[43] Vzhledem k tomu, že AMNH 587 byl prakticky totožný se zachovanou rukou Tanycolagreus exemplář typu, je nyní považován za příslušníka tohoto dinosaura a nikoli za Ornitholestes.[44] Po tomto přeřazení Phil Senter (2006) poznamenal, že „naše znalosti o Ornitholestes lze nyní čerpat pouze z holotypu. “[45] John Foster (2007) uvedl, že některé fragmenty z Suchý lom Mesa může patřit Ornitholestes,[46] ačkoli tyto ještě nebyly popsány.
V roce 1920 Charles Whitney Gilmore dospěl k závěru, že Ornitholestes byl totožný s Coelurus;[47] v roce 1934 Oliver Perry Hay rozpoznal pouze rozdíl na úrovni druhů, pojmenování a Coelurus hermanni,[48] ale v roce 1980 John Ostrom oživil rod.[49]
Klasifikace
The infrařádek Coelurosauria, vytvořený v roce 1914 autorem Friedrich von Huene, byl tradičně a taxonomický odpadkový koš do kterého byly umístěny všechny malé teropody.[50] Ornitholestes, kvůli jeho malé velikosti, byl proto obecně klasifikován jako coelurosaur.[51] V roce 1986 Jacques Gauthier předefinoval tento a několik dalších paleontologických termínů přísnějším způsobem na základě kladistický metody. Tetanurae byl definován jako moderní ptáci a všechny theropody více spjaté s moderními ptáky než s ceratosaury zatímco Coelurosauria nyní zahrnovala všechny členy Tetanurae, které jsou více spjaty s moderními ptáky než s nimi carnosaurs.[52] V roce 1988 Gregory S. Paul navrhl to Ornitholestes byl velmi podobný ve struktuře lebky Proceratosaurus, Middle Jurassic theropod z Anglie.[5] Umístil tyto dva rody dohromady do Ornitholestinae - nové podčeledi pod Allosauridae —A spekulovali, že mají užší příbuznost s mnohem většími Allosaurus než jiným malým teropodům.[53] Nicméně klasifikace Ornitholestes a Proceratosaurus protože alosaurští příbuzní se ukázali jako neudržitelní (ten se od té doby ukázal jako tyrannosauroid ) a Paul to nakonec opustil.[54] Ukázaly se všechny publikované kladistické analýzy Ornitholestes být coelurosaurem, jak ho definoval Gauthier. Některé analýzy prokázaly podporu hypotézy, že jde o nejprimitivnějšího člena skupiny Maniraptora, ačkoli důkladnější analýzy naznačují, že je to primitivnější než Maniraptoriformes, a případně blízký příbuzný „compsognathid“ Juravenator starkii.[55]
Následující rodokmen ilustruje syntézu vztahů hlavních skupin coelurosaurianů na základě různých studií provedených v roce 2010.[56]
| Coelurosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||


Paleobiologie
Ve studii z roku 2001, kterou provedl Bruce Rothschild a další paleontologové, bylo zmíněno 20 kostí nohou Ornitholestes byly vyšetřeny na známky stresová zlomenina, ale nikdo nebyl nalezen.[57]
Funkce přední končetiny
Biomechanická studie provedená Philem Senterem (2006) použila kloubové odlitky Ornitholestes zadejte pravou přední končetinu vzorku a určete jeho rozsah pohybu.[58] Senter zjistil, že antebrachium (předloktí) se může volně houpat do 95 let° rozsah.[59] Když ohnutý (ohnutý dovnitř) v loketním kloubu v maximální možné míře, humerus (horní část paže) a poloměr (dolní část paže) tvořily úhel 53 °.[59] Schopnost Ornitholestes pro Maniraptoriformes je charakteristické ohýbat předloktí na úhel výrazně ostřejší než 90 °, ale chybí u primitivnějších teropodů, jako je Koelophysis a Allosaurus.[60]
I když plně prodloužena (narovnal se) v lokti, předloktí nevytvořilo a rovný úhel, nedosahující tohoto o 22 °.[59] Pronace (zkroucení, aby dlaňová strana ruky směřovala dolů) předloktí bylo nemožné, protože v poloměru a ulně chyběly valivé plochy, což znamená, že předloktí bylo ve stálém stavu supinace.[24]
Když Ornitholestes ohnuté lokty, to by způsobilo, že se předloktí posunou dovnitř, k jeho středové linii.[61] Mohlo to využít této schopnosti uchopit kořist oběma rukama současně.[62]
Strava

Henry Fairfield Osborn, v jeho popisu 1903 Ornitholestes, zaznamenal jeho velké a kuželovité přední zuby, „rychlou sílu uchopení“ jeho ruky a „vyvažovací sílu“ jeho ocasu; tyto interpretoval jako adaptace na kořist současných ptáků.[31] Osborn později tuto hypotézu zavrhl, což v roce 1917 naznačil Ornitholestes představil raná stadia přechodu od masožravého k více býložravému životnímu stylu, ale ne dříve Charles R. Knight nakreslil vlivnou a široce publikovanou ilustraci Ornitholestes honí Archeopteryx.[63] Rytířská ilustrace a další z ní odvozené se i nadále objevovaly v knihách dinosaurů zaměřených na populární publikum po celé 20. století.[51]
David Norman (1985) připustil, že je to „jen možné, i když není příliš pravděpodobné“ Ornitholestes možná chytili a snědli primitivní ptáky.[64] Nejnovější autoři však místo toho navrhli dietu malých suchozemských obratlovců. Savci, ještěrky, žáby, mloci, rhynchocephalians a mláďata dinosaurů by všichni byli potenciální kořist.[65] Gregory S. Paul (1988) si to myslel Ornitholestes mohl použít své kuželovité přední zuby k chytání ryb.[13] Norman (1985) navrhl, že to mohutná lebka a čelisti mohly dovolit Ornitholestes řešit „větší a aktivnější kořist“ než jiné malé teropody.[66] V tomto duchu to spekuloval David Lambert (1993) Ornitholestes, kdyby to byl lovec smeček, mohl by být schopen „se ujmout ornitopody velký jako napůl dospělý Camptosaurus."[67]
V roce 2007 John Foster, specialista na formaci Morrison, navrhl možnost rozdělení oddílů mezi Ornitholestes a jeho současník Coelurus, který byl ve stejném rozsahu velikostí.[68] Teorizoval to s velkýma očima Ornitholestes mohl být specializovaný na noční lov, zatímco Coelurus se možná zaměřili na ty druhy kořisti, které byly aktivní během dne.[68] Foster však poznamenal, že tato hypotéza byla do značné míry spekulativní; nedostatek dochovaného materiálu lebky z Coelurus znemožňuje ověřit, zda jeho oči byly proporcionálně menší než oči Ornitholestes.[69]
Paul (1988) odhaduje, že 12 kg (26 lb) endotermický Ornitholestes by měl denní stravovací požadavek asi 700 g (1,54 lb) masa.[70]
Peří
Ornitolog Percy Lowe předpokládal v roce 1944, že Ornitholestes mohl nést peří.[71] Tato interpretace však byla po více než půl století do značné míry ignorována; většina rekonstrukcí teropodních dinosaurů, včetně Ornitholestes, vylíčil je s plazí šupinatou kůží. Jednou z mála výjimek byla Gregory S. Paul Draví dinosauři světa (1988).[72] Robert T. Bakker je Kacířství o dinosaurech (1986) přesně předpovídali přítomnost peří na dromaeosauridech a tvrdili, že všichni dinosauři byli endotermní, přesto nezahrnuli peří do své ilustrace Ornitholestes.[73]
V roce 1996 primitivní coelurosaur Sinosauropteryx byl objeven v Číně. Zachovaná fosilie ukázala, že tento dinosaurus nesl plášť rudimentárního kožešinového peří. V důsledku tohoto a dalších podobných objevů většina paleontologů nyní považuje za pravděpodobné, že všichni coelurosaurové měli nějaká izolační peří, nebo pocházeli od předků, kteří to měli.[74] John Foster (2007) na základě své fylogenetické polohy vyvodil, že peří z Ornitholestes pravděpodobně by byl primitivnější než ptáci; pravděpodobně „by pokryly tělo kromě nohou v krátké srsti, možná delšími peřími lemujícími horní část lebky nebo krku a zadní okraj předloktí.“[75] Za předpokladu, že by byly přítomny, bylo by toto peří použito k izolaci a případně také k izolaci napjatá vejce.[75]
Reference
- ^ Turner, C.E. a Peterson, F., (1999). „Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, USA“ Str. 77–114 v Gillette, D.D. (vyd.), Paleontologie obratlovců v Utahu. Geologická služba v Utahu Různé Publikace 99-1.
- ^ Lambert (1993), str. 79
- ^ Paul (1988A), s. 306; Norman (1985), str. 42; Norman (1990), str. 293
- ^ Osborn (1903), str. 460; Chure (1998), str. 238
- ^ A b Paul (1988b), s. 6
- ^ Paul (1988b), s. 3; Norman (1990), str. 293
- ^ Osborn (1903), str. 460; Osborn (1917), str. 734
- ^ Paul (1988b), s. 4; Norman (1990), str. 293
- ^ Osborn (1903), str. 460
- ^ Paul (1988b), s. 3; Norman (1990), str. 292
- ^ Cristiano dal Sasso & Simone Maganuco, 2011, Scipionyx samniticus (Theropoda: Compsognathidae) ze spodní křídy Itálie - Osteologie, ontogenetické hodnocení, fylogeneze, anatomie měkkých tkání, taphonomy a paleobiologie, Memorie della Società Italiana de Scienze Naturali e del Museo Civico di Storia Naturale di Milano XXXVII(I): 1-281
- ^ Osborn (1903), str. 460; Norman (1990), str. 293
- ^ A b Paul (1988A), s. 303
- ^ Rauhut (2003), str. 27; Tesař et al. (2005A), s. 53
- ^ Paul (2010), str. 123
- ^ Tesař et al. (2005A), str. 49, 54
- ^ Osborn (1917), str. 734; Lambert (1993), str. 78
- ^ Osborn (1917), str. 735–736
- ^ Tesař et al. (2005A), s. 58
- ^ Tesař et al. (2005A), s. 49, 69
- ^ Ostrom (1969), str. 146; Paul (2002), str. 236
- ^ Osborn (1917), str. 738; Tesař et al. (2005b), s. 45
- ^ Tesař et al. (2005A), s. 60, 62
- ^ A b C Senter (2006), s. 1030
- ^ Paul (1988A), s. 306
- ^ Osborn (1917), str. 737
- ^ Tesař et al. (2005A), s. 67
- ^ Osborn (1903), str. 463
- ^ Ostrom (1969), str. 161
- ^ Ostrom (1969), str. 161; Paul (1988b), s. 6
- ^ A b C d Osborn (1903), str. 459
- ^ Osborn (1917), str. 735; Paul (1988b), s. 3
- ^ Norman (1985), str. 39; Foster (2007), s. 178
- ^ Paul (1988A), s. 305
- ^ Paul (1988A), s. 305; Foster (2007), s. 178
- ^ Long a Schouten (2008), s. 73
- ^ Fastovsky a Weishampel (2005), s. 291
- ^ Paul D. Brinkman, 2010, The Second Jurassic Dinosaur Rush: Museums & Paleontology in America at the Turn of the Twentieth Century. University of Chicago Press, 345 s., ISBN 978-0-226-07472-6
- ^ Osborn (1903), str. 459–464
- ^ Osborn (1903), str. 459; Osborn (1917), str. 733
- ^ Osborn (1903), str. 459; Osborn (1917), str. 736
- ^ Paul (1988b), s. 3
- ^ Tesař et al. (2005b), s. 23, 39
- ^ Tesař et al. (2005b), s. 39
- ^ Senter (2006), s. 1029
- ^ Foster (2007), s. 78, 179
- ^ Gilmore, C.W., 1920, „Osteologie masožravé dinosaurie v Národním muzeu Spojených států, se zvláštním zřetelem na rody Antrodemus (Allosaurus) a Ceratosaurus“, Bulletin Národní muzeum USA CX: 1-154
- ^ Hay, O.P., 1930, Druhá bibliografie a katalog fosilních obratlovců Severní Ameriky. Carnegie Institution of Washington. 390 (II): 1-1074
- ^ Ostrom (1980)
- ^ Paul (1988A), s. 188–189; Norman (1990), str. 280
- ^ A b Glut (1997), str. 645
- ^ Holtz et al. (2004), str. 71
- ^ Paul (1988A), s. 302; Paul (1988b), s. 3, 6
- ^ Paul (2010), str. 124
- ^ Godefroit, Pascal; Cau, Andrea; Hu, Dong-Yu; Escuillié, François; Wu, Wenhao; Dyke, Gareth (2013). „Jurský avialanský dinosaurus z Číny řeší ranou fylogenetickou historii ptáků“. Příroda. 498 (7454): 359–362. Bibcode:2013 Natur.498..359G. doi:10.1038 / příroda12168. PMID 23719374.
- ^ Hendrickx, C., Hartman, S.A., & Mateus, O. (2015). Přehled objevů a klasifikace jiných než ptačích teropodů. PalArch's Journal of Vertebrate Palaeontology, 12(1): 1-73.
- ^ Rothschild, B., Tanke, DH a Ford, TL, 2001, Theropod stresové zlomeniny a avulze šlachy jako vodítko k aktivitě: In: Mesozoic Vertebrate Life, edited by Tanke, DH, and Carpenter, K., Indiana University Press, p. 331-336.
- ^ Senter (2006), s. 1029–1030
- ^ A b C Senter (2006), s. 1030–1031
- ^ Senter (2006), s. 1032
- ^ Senter (2006), s. 1030–1032
- ^ Senter (2006), s. 1031
- ^ Osborn (1917), str. 733–734; Glut (1997), str. 645
- ^ Norman (1985), str. 39
- ^ Norman (1985), str. 39; Paul (1988), str. 306; Foster (2007), s. 179; Long a Schouten (2008), s. 73
- ^ Norman (1985), str. 42
- ^ Lambert (1993), str. 78
- ^ A b Foster (2007), s. 180
- ^ Foster (2007), s. 180–181
- ^ Paul (1988A), s. 404
- ^ Witmer (1992), str. 444
- ^ Paul (1988A), s. 122–123, 211
- ^ Bakker (1986), str. 99, 310
- ^ Chatterjee a Templin (2004), str. 277; Fastovsky a Weishampel (2005), s. 329–330; Currie (2005), str. 368
- ^ A b Foster (2007), s. 179
Zdroje
- Osborn, Henry Fairfield (1903). "Ornitholestes hermanni, nový komsognathoidní dinosaurus z horní jury “. Bulletin of the American Museum of Natural History. 19 (12): 459–464. hdl:2246/1502.
- Osborn, Henry Fairfield (1917). "Skeletální adaptace Ornitholestes, Struthiomimus, Tyranosaurus". Bulletin of the American Museum of Natural History. 35 (43): 733–771. hdl:2246/1334.
- Ostrom, John H. (1969). "Osteologie Deinonychus antirrhopus, neobvyklý teropod ze spodní křídy v Montaně “. Peabody Museum of Natural History Bulletin. 30: 1–165.
- Ostrom, John H. (1980). "Coelurus a Ornitholestes: Jsou stejní? ". V Jacobs, Louis L. (ed.). Aspekty historie obratlovců: Pokusy o čest Edwina Harrisa Colberta. Museum of Northern Arizona Press. str. 245–256. ISBN 978-0-89734-052-6.
- Norman, David B. (1985). „Coelurosaurs“. Ilustrovaná encyklopedie dinosaurů. Salamander Books Ltd. str. 38–43. ISBN 978-0-517-46890-6.
- Paul, Gregory S. (1988A). „Ornitholestians and Allosaurs“. Draví dinosauři světa. Simon & Schuster. str.302–318. ISBN 978-0-671-61946-6. Zkontrolujte hodnoty data v:
| rok =(Pomoc) - Paul, Gregory S. (1988b). „Malí draví dinosauři ve střední druhohorě: rohatí teropody Morrisona a Great Oolite -Ornitholestes a Proceratosaurus—A srpkovité drápy teropodů Cloverly, Djadokhta a Judith River -Deinonychus, Velociraptor a Saurornitholestes". Hunteria. 2 (4): 1–9. Zkontrolujte hodnoty data v:
| rok =(Pomoc) - Norman, David B. (1990). „Problematic Theropoda: Coelurosaurs“. v Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). Dinosaurie. University of California Press. str. 280–305. ISBN 978-0-520-06726-4.
- Lambert, David (1993). "Ornitholestes". The Ultimate Dinosaur Book. Dorling Kindersley. str.78–79. ISBN 978-1-56458-304-8.
- Glut, Donald F. (1997). "Ornitholestes". Dinosauři: Encyklopedie. McFarland & Company. 643–646. ISBN 978-0-89950-917-4.
- Chure, Daniel (1998). „Na oběžné dráze dinosaurů z Theropodu“ (PDF). Gaia (18): 233–240. Archivovány od originál (PDF) dne 19. 7. 2011. Citováno 2011-02-06.
- Paul, Gregory S. (2002). „Byli někteří dinosauři také ptáky bez světla?“. Dinosaurs of the Air: Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press. str.224–257. ISBN 978-0-8018-6763-7.
- Rauhut, Oliver W.M. (2003). "Provozní taxonomické jednotky". Vzájemné vztahy a vývoj bazálních teropodních dinosaurů. Paleontologická asociace. s. 12–43. ISBN 978-0-901702-79-1.
- Holtz, Thomas R.; Molnár, Ralph E.; Currie, Philip J. (2004). „Bazální tetanura“. v Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria: Second Edition. University of California Press. 71–110. ISBN 978-0-520-24209-8.CS1 maint: více jmen: seznam autorů (odkaz)
- Fastovský, David E .; Weishampel, David B. (2005). „Theropoda I: Příroda červená v zubu a drápu“. Evoluce a zánik dinosaurů. Cambridge University Press. 265–299. ISBN 978-0-521-81172-9.
- Carpenter, Kenneth; Miles, Clifford; Ostrom, John H.; Cloward, Karen (2005A). „Redescription of Small Maniraptoran Theropods Ornitholestes a Coelurus". V Carpenter, Kenneth (ed.). Masožraví dinosauři. Život minulosti. Indiana University Press. str. 49–71. ISBN 978-0-253-34539-4. Zkontrolujte hodnoty data v:
| rok =(Pomoc) - Carpenter, Kenneth; Miles, Clifford; Cloward, Karen (2005b). „Nový malý teropod z vrchní jury Morrisonské formace ve Wyomingu“. V Carpenter, Kenneth (ed.). Masožraví dinosauři. Život minulosti. Indiana University Press. 23–48. ISBN 978-0-253-34539-4. Zkontrolujte hodnoty data v:
| rok =(Pomoc) - Senter, Phil (2006). "Funkce předních končetin v Ornitholestes hermanni Osborn (Dinosauria, Theropoda) ". Paleontologie. 49 (5): 1029–1034. doi:10.1111 / j.1475-4983.2006.00585.x.
- Foster, John (2007). „Gargantuan minuskule: Morrisonský zvěřinec, část II“. Jurský západ: Dinosauři formace Morrison a jejich svět. Indiana University Press. 162–242. ISBN 978-0-253-34870-8.
- Long, John A.; Schouten, Peter (2008). "Ornitholestes a příbuzní". Feathered Dinosaurs: Původ ptáků. Oxford University Press. 72–77. ISBN 978-0-19-537266-3.
- Paul, Gregory S. (2010). "Ornitholestes hermanni". The Princeton Field Guide to Dinosaurs. Princeton University Press. str.123–124. ISBN 978-0-691-13720-9.













