GADD45G - GADD45G




Zastavení růstu a gama GADD45 indukovatelné proteinem poškození DNA je protein že u lidí je kódován GADD45G gen na chromozom 9. GADD45G je také známý jako CR6, DDIT2, GRP17, OIG37 a GADD45gamma.[5] GADD45G je zapojen do několika různých procesů, včetně sexuálního vývoje,[6] vývoj mozku specifický pro člověka,[7] potlačení nádoru,[8] a buněčná stresová reakce.[9] GADD45G interaguje s několika dalšími proteiny, které se podílejí na opravě DNA, buněčný cyklus řízení, apoptóza, a stárnutí.[6] Nízká exprese GADD45G byla spojena s mnoha typy rakovina.[10]
Dějiny
GADD45G byl původně klonován Beadlingem pod názvem CR6 v roce 1993. V tomto experimentu bylo zaznamenáno, že několik genů včetně GADD45G bylo indukováno IL-2 a byly identifikovány jako geny pro okamžitou odezvu v T lymfocyty.[11] Jeho role jako supresor nádoru byl objeven v roce 1999 Zhangem.[12] To dostalo název OIG37 od Nakayama kvůli jeho regulaci Oncostatin M., u kterého bylo zjištěno, že je schopen inhibovat růst.[13] Nakonec se také stal známým jako protein 17 související s Gadd během jeho izolace z a cDNA knihovna Suzuki kvůli homologie s Gadd45.[5]
Struktura a funkce
GADD45G je členem skupiny genů, jejichž hladiny transkriptů se zvyšují po stresových podmínkách zastavení růstu a léčbě látkami poškozujícími DNA. Protein kódovaný tímto genem reaguje na stresy prostředí zprostředkováním aktivace str /JNK cesta přes MTK1 /MEKK4 kináza.[14] GADD45G je zase regulován proti proudu NF-kB.[8]
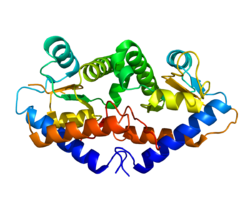
Krystalová struktura GADD45G odhaluje a dimer ze čtyř paralelních šroubovic. Centrální oblast obsahuje vysoce kyselou náplast, kde umožňuje interakci s cdc2, PCNA, a p21. Paralela izoforma GADD45G je aktivní forma.[15]
Tento gen hraje roli v regulaci buněčného cyklu. GADD45G brání kinázové schopnosti cyklinu b1 /Cdk1 komplex takovým způsobem, aby komplex nerozpadl. Hraje roli při aktivaci S a Kontrolní body G2 / M.[16]
V cestě mužského pohlavního vývoje je GADD45G nezbytný pro aktivaci SRY, což vede ke správné tvorbě pohlavní žlázy a určení pohlaví. K tomu může dojít prostřednictvím interakce GADD45G s str Signální dráha MAPK.[6]
Vymazání zesilovač v blízkosti genu GADD45G souvisí se zvýšenou proliferací neuronální buňky, které by mohly odpovídat za část rozdílu v nervovém vývoji mezi lidmi a jinými druhy.[7] Delece zesilovače snižuje expresi genu v přední mozek umožňující větší růst mozku u lidí.[17]
GADD45G je zapojen do zubního lékařství epiteliální proliferace buněk. GADD45G je vyjádřen v smaltované uzly, kde reguluje genovou expresi a růst buněk. Gen moduluje proliferaci epiteliálních buněk zprostředkovanou p21 aktivací dráhy p38 MAPK během vývoje zubů.[18]
Existuje rozdílné vyjádření Xenopus homolog GADD45G v embryonálním vývoji. Hraje velkou roli v nervovém a mozkovém vývoji s GADD45A. GADD45G a GADD45A příklepy souvisí s nevhodným gastrulace, vadný růst hlavy a kratší osy. GADD45G a GADD45A působí nadbytečně k řízení buněčného růstu, umožněte buňkám pohyb pluripotence pomáhání buňky se diferencují.[19]
Paměť
Během učební zkušenosti se skupina genů rychle exprimuje v mozek. Tato indukovaná genová exprese je považována za důležitou pro zpracování naučené informace. Takové geny jsou známé jako okamžité rané geny. V rámci prelimbiky prefrontální kůra, GADD45G Gen je okamžitě exprimován a je vyžadován pro konsolidaci typu učení u myší paměť asociativního strachu.[20] Obecně, genová exprese často může být epigeneticky vyvolané demetylace z 5-methylcytosin s) v promotor genů regionech. Protein GADD45G funguje v oprava poškození DNA. GADD45G může být také zapojen do rozpoznávání 5-methylcytosinu jako změny v DNA, kterou je třeba opravit, aby bylo možné vyvolat geny související s učením. GADD45G tedy může řídit rychlou demetylaci methylcytosinu v promotorových oblastech genů souvisejících s učením pomocí Oprava DNA proces[20](viz také Epigenetika v učení a paměti ).
Interakce
GADD45G provádí mnoho dříve uvedených funkcí s mnoha různými interakcemi. Bylo zjištěno, že GADD45G inhibuje Cdk1 kináza aktivita, která by způsobila narušení buněčného růstu.[16] Interaguje také s CRIF, což způsobuje inhibici Cdc2-cyklinu B1 a Cdk-cyklinu E.[21] GADD45 také pracuje s inhibitorem kinázy závislým na cyklinu p21, což může také způsobit zastavení růstu.[22] Bylo zjištěno, že GADD45G je zapojen do str Cesta MAPK prostřednictvím interakcí s MAP3K4, což může být důležité při určování pohlaví.[23] Kromě toho GADD45G reguluje replikaci a opravu DNA prostřednictvím svých interakcí s PCNA.[13][22]
Distribuce tkání
U lidí je GADD45G nejvíce exprimován v kosterní sval, ledviny a játra. Tento gen má nízkou expresi v srdce, mozek, slezina, plíce a varlata.[8] GADD45G je vysoce exprimován v placentě.[24]
U embryonální myši je Gadd45g exprimován v neurální trubice, lebeční a hřbetní kořenová ganglia a hřbetní střední mozek.[25]
Savčí buňky ledvinové vnitřní dřeně (IM) běžně čelí hypertonickému stresu a odolávají mu. Takový stres způsobuje poškození DNA, na které IM buňky reagují zastavením buněčného cyklu. Všichni tři GADD45 izoformy GADD45A, GADD45B a GADD45G jsou indukovány akutní hypertonicitou v myších IM buňkách. Maximální indukce nastává 16-18 hodin po nástupu hypertonicity. GADD45G je indukován silněji (7krát) než GADD45B (3krát) a GADD45A (2krát). Hypertonicita různých forem (NaCl, KCl, sorbitol nebo manitol) vždy vyvolává GADD45 přepisy, zatímco nehypertonická hyperosmolalita (močovina) nemá žádný účinek. Aktinomycin D nezabrání hypertonické indukci GADD45, což naznačuje, že stabilizace mRNA je mechanismus, který tuto indukci zprostředkovává.[26]
Klinický význam
V mnoha druzích rakovinných buněk je GADD45G regulováno.[10] Existuje nízký výraz kvůli methylace promotoru GADD45G.[18] Tuto nízkou expresi lze také vysvětlit zvýšenou aktivací NF-kB.[27]
Methylace GADD45G je vidět u mnoha druhů rakoviny. v rakovina jícnu úroveň exprese a stav methylace genu se podílejí na prognóze jícnu spinocelulární karcinom. Demetylace genu může mít některé příznivé účinky.[18] Podobné okolnosti jsou vidět v žaludeční kardioadenokarcinomy kde je GADD45G umlčen.[28] Úrovně methylace GADD45G se také měří v diagnostice rakoviny pankreatu a kolorektálního karcinomu.[29]
V hypofýza „GADD45G je látka potlačující růst. U mnoha rakovinových mas hypofýzy dochází ke ztrátě exprese genu.[30] Gen hraje roli v rakovina prostaty jako supresor nádoru. V těchto rakovinných buňkách Vitamín D může vyvolat expresi GADD45G.[31] GADD45G by mohl být terčem terapeutického přínosu pro rakovinu prostaty.[32]
v rakovinné jaterní buňky, GADD45G je regulován dolů. Podílí se na negativní regulaci signální dráhy Jak-Stat3. Působí jako supresor nádoru HCC buňky podporou buněčné smrti nebo zastavení růstu. Když je exprese GADD45G nízká, jaterní buňky mohou být schopny obejít fázi zastavení růstu, což vede k rakovinným buňkám.[10]
Přítomnost GADD45G v močový systém souvisí také s nemoc ledvin. Byly poškozeny renální buňky exprimující gen.[33]
Upregulace Gadd45g způsobená hormony může odpovídat za změny v děloze myši.[34]
Viz také
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000130222 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000021453 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b Suzuki M, Watanabe TK, Fujiwara T, Takahashi E, Tanigami A (říjen 1999). „Molekulární klonování, exprese a mapování nové lidské cDNA, GRP17, vysoce homologní s lidským gadd45 a myším MyD118“. J Hum Genet. 44 (5): 300–3. doi:10,1007 / s100380050164. PMID 10496071.
- ^ A b C Johnen H, González-Silva L, Carramolino L, Flores JM, Torres M, Salvador JM (2013). „Gadd45g je nezbytný pro primární určení pohlaví, mužskou plodnost a vývoj varlat“. PLOS ONE. 8 (3): e58751. doi:10.1371 / journal.pone.0058751. PMC 3596291. PMID 23516551.
- ^ A b McLean CY, Reno PL, Pollen AA, Bassan AI, Capellini TD, Guenther C, Indjeian VB, Lim X, Menke DB, Schaar BT, Wenger AM, Bejerano G, Kingsley DM (březen 2011). „Ztráta regulační DNA specifické pro člověka a vývoj vlastností specifických pro člověka“. Příroda. 471 (7337): 216–9. doi:10.1038 / nature09774. PMC 3071156. PMID 21390129.
- ^ A b C Tamura RE, de Vasconcellos JF, Sarkar D, Libermann TA, Fisher PB, Zerbini LF (červen 2012). „GADD45 proteiny: centrální hráči v tumorigenezi“. Curr. Mol. Med. 12 (5): 634–51. doi:10.2174/156652412800619978. PMC 3797964. PMID 22515981.
- ^ Liebermann DA, Hoffman B (2007). „Gadd45 v reakci hematopoetických buněk na genotoxický stres“. Blood Cells Mol. Dis. 39 (3): 329–35. doi:10.1016 / j.bcmd.2007.06.006. PMC 3268059. PMID 17659913.
- ^ A b C Zhang L, Yang Z, Ma A, Qu Y, Xia S, Xu D, Ge C, Qiu B, Xia Q, Li J, Liu Y (leden 2014). „Zastavení růstu a poškození DNA 45G down-regulace přispívá k Janus kinase / signálnímu převodníku a aktivátoru aktivace transkripce 3 a úniku buněčného stárnutí v hepatocelulárním karcinomu“. Hepatologie. 59 (1): 178–89. doi:10.1002 / hep.26628. PMID 23897841. S2CID 39582166.
- ^ Beadling C, Johnson KW, Smith KA (duben 1993). „Izolace okamžitých a časných genů indukovaných interleukinem 2“. Proc. Natl. Acad. Sci. USA. 90 (7): 2719–23. doi:10.1073 / pnas.90.7.2719. PMC 46167. PMID 7681987.
- ^ Zhang W, Bae I, Krishnaraju K, Azam N, Fan W, Smith K, Hoffman B, Liebermann DA (září 1999). „CR6: Třetí člen rodiny genů MyD118 a Gadd45, který funguje při negativní kontrole růstu“. Onkogen. 18 (35): 4899–907. doi:10.1038 / sj.onc.1202885. PMID 10490824.
- ^ A b Nakayama K, Hara T, Hibi M, Hirano T, Miyajima A (srpen 1999). „Nový gen indukující onkostatin M OIG37 tvoří genovou rodinu s MyD118 a GADD45 a negativně reguluje buněčný růst.“. J. Biol. Chem. 274 (35): 24766–72. doi:10.1074 / jbc.274.35.24766. PMID 10455148.
- ^ Takekawa M, Saito H (prosinec 1998). "Rodina stresově indukovatelných proteinů podobných GADD45 zprostředkovává aktivaci stresově citlivých MTK1 / MEKK4 MAPKKK". Buňka. 95 (4): 521–30. doi:10.1016 / S0092-8674 (00) 81619-0. PMID 9827804. S2CID 18980341.
- ^ Zhang W, Fu S, Liu X, Zhao X, Zhang W, Peng W, Wu C, Li Y, Li X, Bartlam M, Zeng ZH, Zhan Q, Rao Z (2011). „Krystalová struktura lidského Gadd45γ [opraveno] odhaluje aktivní dimer“. Proteinová buňka. 2 (10): 814–26. doi:10.1007 / s13238-011-1090-6. PMC 4875293. PMID 22058036.
- ^ A b Vairapandi M, Balliet AG, Hoffman B, Liebermann DA (2002). „GADD45b a GADD45g jsou inhibitory kinázy cdc2 / cyklinB1 s rolí v kontrolních bodech buněčného cyklu S a G2 / M vyvolaných genotoxickým stresem“. J. Cell. Physiol. 192 (3): 327–38. doi:10.1002 / jcp.10140. PMID 12124778. S2CID 19138273.
- ^ Iskow RC, Gokcumen O, Lee C (2012). „Zkoumání role variant počtu kopií v lidské adaptaci“. Trendy v genetice. 28 (6): 245–257. doi:10.1016 / j.tig.2012.03.002. ISSN 0168-9525. PMC 3533238. PMID 22483647.
- ^ A b C Ishida K, Yuge Y, Hanaoka M, Yasukawa M, Minami Y, Ogawa M, Masumoto KH, Shigeyoshi Y, Saito M, Tsuji T (srpen 2013). "Gadd45g reguluje proliferaci buněk zubního epitelu prostřednictvím exprese p21 MAPK zprostředkované p21". Geny buňky. 18 (8): 660–71. doi:10.1111 / gtc.12067. PMID 23751077. S2CID 23664498.
- ^ Kaufmann LT, Niehrs C (2011). „Gadd45a a Gadd45g regulují vývoj nervů a opuštění pluripotence v Xenopus“. Mechanismy rozvoje. 128 (7–10): 401–411. doi:10.1016 / j.mod.2011.08.002. ISSN 0925-4773. PMID 21854844.
- ^ A b Li X, Marshall PR, Leighton LJ, Zajaczkowski EL, Wang Z, Madugalle SU, Yin J, Bredy TW, Wei W. Protein Gadd45γ související s opravou DNA reguluje dočasné kódování okamžitého časného projevu genu v prelimbické prefrontální kůře a je Vyžadováno pro konsolidaci asociativní strachu z paměti. J. Neurosci. 2019 6. února; 39 (6): 970-983. doi: 10.1523 / JNEUROSCI.2024-18.2018. Epub 2018 12. prosince. Erratum in: J Neurosci. 2019 15. května; 39 (20): 3993. PMID: 30545945
- ^ Chung HK, Yi YW, Jung NC, Kim D, Suh JM, Kim H, Park KC, Song JH, Kim DW, Hwang ES, Yoon SH, Bae YS, Kim JM, Bae I, Shong M (červenec 2003). „Faktor 1 interagující s CR6 interaguje s proteiny rodiny Gadd45 a moduluje buněčný cyklus“. J. Biol. Chem. 278 (30): 28079–88. doi:10,1074 / jbc.M212835200. PMID 12716909.
- ^ A b Azam N, Vairapandi M, Zhang W, Hoffman B, Liebermann DA (leden 2001). „Interakce CR6 (GADD45gamma) s proliferujícím buněčným jaderným antigenem brání negativní kontrole růstu“. J. Biol. Chem. 276 (4): 2766–74. doi:10,1074 / jbc.M005626200. PMID 11022036.
- ^ Warr N, Carre GA, Siggers P, Faleato JV, Brixey R, Pope M, Bogani D, Childers M, Wells S, Scudamore CL, Tedesco M, del Barco Barrantes I, Nebreda AR, Trainor PA, Greenfield A (listopad 2012) . „Interakce Gadd45γ a Map3k4 regulují stanovení myšího varlete prostřednictvím p38 MAPK zprostředkované kontroly Sryho exprese“. Dev. Buňka. 23 (5): 1020–31. doi:10.1016 / j.devcel.2012.09.016. PMC 3526779. PMID 23102580.
- ^ „Entrez Gene: GADD45G zastavování růstu a indukovatelné poškození DNA, gama“.
- ^ Kaufmann LT, Gierl MS, Niehrs C (2011). "Exprese Gadd45a, Gadd45b a Gadd45g během embryonálního vývoje myší". Vzory genového výrazu. 11 (8): 465–470. doi:10.1016 / j.gep.2011.07.005. ISSN 1567-133X. PMID 21843656.
- ^ Chakravarty D, Cai Q, Ferraris JD, Michea L, Burg MB, Kültz D (listopad 2002). „Tři izoformy GADD45 přispívají k fenotypu hypertonického stresu vnitřních dřeňových buněk myší“. American Journal of Physiology. Fyziologie ledvin. 283 (5): F1020–9. doi:10.1152 / ajprenal.00118.2002. PMID 12372778.
- ^ Liebermann DA, Tront JS, Sha X, Mukherjee K, Mohamed-Hadley A, Hoffman B (2011). „Senzory stresu Gadd45 u malignity a leukémie“. Crit Rev Oncog. 16 (1–2): 129–40. doi:10.1615 / critrevoncog.v16.i1-2.120. PMC 3268054. PMID 22150313.
- ^ Guo W, Dong Z, Guo Y, Chen Z, Kuang G, Yang Z (2013). "Methylace zprostředkovaná represi exprese GADD45A a GADD45G v adenokarcinomu žaludku". International Journal of Cancer. 133 (9): 2043–2053. doi:10.1002 / ijc.28223. ISSN 0020-7136. PMID 23616123. S2CID 36324243.
- ^ Zhang W, Li T, Shao Y, Zhang C, Wu Q, Yang H, Zhang J, Guan M, Yu B, Wan J (srpen 2010). „Semi-kvantitativní detekce hladin methylace GADD45-gama u rakoviny žaludku, kolorektálního karcinomu a rakoviny slinivky břišní s použitím metylačně citlivé analýzy tání s vysokým rozlišením“. J. Cancer Res. Clin. Oncol. 136 (8): 1267–73. doi:10.1007 / s00432-010-0777-z. PMID 20111973. S2CID 37780631.
- ^ Zhang X, Sun H, Danila DC, Johnson SR, Zhou Y, Swearingen B, Klibanski A (2002). „Ztráta exprese genu GADD45 gama, genu inhibujícího růst, v adenomech lidské hypofýzy: důsledky pro tumorigenezi“. J Clin Endocrinol Metab. 87 (3): 1262–7. doi:10.1210 / jcem.87.3.8315. PMID 11889197.
- ^ Flores O, Burnstein KL (2010). „GADD45gamma: nový gen regulovaný vitaminem D, který je antiproliferativní v buňkách rakoviny prostaty“. Endokrinologie. 151 (10): 4654–64. doi:10.1210 / cs.2010-0434. PMC 2946153. PMID 20739400.
- ^ Liebermann DA, Hoffman B (říjen 2011). „Rakovina prostaty: JunD, Gadd45a a Gadd45g jako terapeutické cíle“. Buněčný cyklus. 10 (20): 3428. doi:10,4161 / cc.10.20.17528. PMID 22030693.
- ^ Yu S, Cho J, Park I, Kim SJ, Kim H, Shin GT (2009). "Exprese GADD45gamma v moči je spojena s progresí lgA nefropatie". Jsem J. Nephrol. 30 (2): 135–9. doi:10.1159/000209317. PMID 19293565. S2CID 46326535.
- ^ Ivanga M, Labrie Y, Calvo E, Belleau P, Martel C, Pelletier G, Morissette J, Labrie F, Durocher F (2009). "Jemná časová analýza transkripční modulace DHT signálních drah ATM / Gadd45g v děloze myši". Molekulární reprodukce a vývoj. 76 (3): 278–288. doi:10,1002 / mrd.20949. ISSN 1040-452X. PMID 18671277. S2CID 9149501.
Další čtení
- Fan W, Richter G, Cereseto A, Beadling C, Smith KA (2000). „Gen 6 pro odpověď na cytokiny indukuje p21 a reguluje růst a zástavu buněk“. Onkogen. 18 (47): 6573–82. doi:10.1038 / sj.onc.1203054. PMID 10597261.
- Gong R, Yu L, Zhang H, Tu Q, Zhao Y, Yang J, Xu Y, Zhao S (2000). "Přiřazení lidského GADD45G k chromozomu 9q22.1 → q22.3 radiačním hybridním mapováním". Cytogenet. Cell Genet. 88 (1–2): 95–6. doi:10.1159/000015496. PMID 10773677. S2CID 45495955.
- Yi YW, Kim D, Jung N, Hong SS, Lee HS, Bae I (2000). „Proteiny rodiny Gadd45 jsou koaktivátory jaderných hormonálních receptorů“. Biochem. Biophys. Res. Commun. 272 (1): 193–8. doi:10,1006 / bbrc.2000.2760. PMID 10872826.
- Yang Q, Manicone A, Coursen JD, Linke SP, Nagashima M, Forgues M, Wang XW (2001). „Identifikace funkční domény v kontrolním bodě G2 / M zprostředkovaném GADD45“. J. Biol. Chem. 275 (47): 36892–8. doi:10,1074 / jbc.M005319200. PMID 10973963.
- Wan Y, Wang Z, Shao Y, Xu Y, Voorhees J, Fisher G (2001). „Exprese GADD45 indukovaná UV zářením je zprostředkována cestou citlivou na oxidanty v kultivovaných lidských keratinocytech a v lidské kůži in vivo“. Int. J. Mol. Med. 6 (6): 683–8. doi:10,3892 / ijmm.6.6.683. PMID 11078829.
- Kovalsky O, Lung FD, Roller PP, Fornace AJ (2001). „Oligomerizace lidského proteinu Gadd45a“. J. Biol. Chem. 276 (42): 39330–9. doi:10,1074 / jbc.M105115200. PMID 11498536.
- Sun L, Gong R, Wan B, Huang X, Wu C, Zhang X, Zhao S, Yu L (2004). „GADD45gamma, down-regulovaná u 65% hepatocelulárního karcinomu (HCC) od 23 čínských pacientů, inhibuje buněčný růst a indukuje zástavu G2 / M buněčného cyklu pro buněčné linie hepatomu Hep-G2“. Mol. Biol. Rep. 30 (4): 249–53. doi:10.1023 / A: 1026370726763. PMID 14672412. S2CID 33065335.
- Jiang F, Wang Z (2004). „Gadd45gamma reaguje na androgeny a inhibuje růst v buňkách rakoviny prostaty“. Mol. Buňka. Endokrinol. 213 (2): 121–9. doi:10.1016 / j.mce.2003.10.050. PMID 15062559. S2CID 54408868.
- Zerbini LF, Wang Y, Czibere A, Correa RG, Cho JY, Ijiri K, Wei W, Joseph M, Gu X, Grall F, Goldring MB, Zhou JR, Libermann TA, Zhou JR (2004). „NF-kappa B zprostředkovaná represe proteinů 45alfa a gama indukovaných zastavením růstu a poškození DNA je nezbytná pro přežití rakovinných buněk“. Proc. Natl. Acad. Sci. USA. 101 (37): 13618–23. doi:10.1073 / pnas.0402069101. PMC 518803. PMID 15353598.
- Goehler H, Lalowski M, Stelzl U, Waelter S, Stroedicke M, Worm U, Droege A, Lindenberg KS, Knoblich M, Haenig C, Herbst M, Suopanki J, Scherzinger E, Abraham C, Bauer B, Hasenbank R, Fritzsche A Ludewig AH, Büssow K, Buessow K, Coleman SH, Gutekunst CA, Landwehrmeyer BG, Lehrach H, Wanker EE (2004). „Síť proteinových interakcí spojuje GIT1, zesilovač agregace huntingtinu, s Huntingtonovou chorobou“. Mol. Buňka. 15 (6): 853–65. doi:10.1016 / j.molcel.2004.09.016. PMID 15383276.