Systém toxin-antitoxin SymE-SymR - SymE-SymR toxin-antitoxin system
| SymR | |
|---|---|
 Zachovaná sekundární struktura SymR RNA. | |
| Identifikátory | |
| Symbol | SymR |
| Rfam | RF01809 |
| Další údaje | |
| RNA typ | Antisense RNA |
| Domény | E-coli |
| PDB struktur | PDBe |
| Toxin SymE typu toxin-antitoxinový systém typu I. | |||||||||
|---|---|---|---|---|---|---|---|---|---|
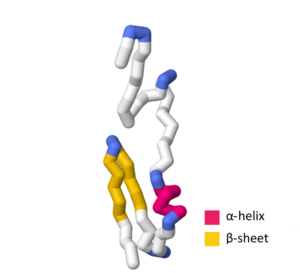 Toxin SymE typu toxin-antitoxinový systém typu I. | |||||||||
| Identifikátory | |||||||||
| Symbol | SymE_toxin | ||||||||
| Pfam | PF13957 | ||||||||
| InterPro | IPR020883 | ||||||||
| STRÁNKA | PS51740 | ||||||||
| |||||||||
| https://swissmodel.expasy.org/repository/uniprot/P39394 | |||||||||
The Systém toxin-antitoxin SymE-SymR sestává z malého symbiotika endonukleáza toxin, SymE a a nekódující RNA symbiotický RNA antitoxin, SymR, který inhibuje translaci SymE.[1] SymE-SymR je toxin-antitoxinový systém typu I a je regulován antitoxinem, SymR.[2] Předpokládá se, že komplex SymE-SymR hraje důležitou roli při recyklaci poškozených RNA a DNA.[1] Vztah a odpovídající struktury SymE a SymR poskytují vhled do mechanismu toxicity a celkové role v prokaryotických systémech.
Objev
SymR byl původně označen RyjC a je to 77 nukleotid (nt) RNA s a σ70 promotér. Bylo zjištěno, že RyjC překrývá yjiW otevřený čtecí rámec na opačném řetězci o 6 nt a byla charakterizována jako antisense RNA který vázal 5 'nepřekládaná oblast z yjiW.[3] Další studie vedla k přejmenování obou yjiW a RyjC na SymE (gen yjiW indukovaný SOS s podobností s Bludiště ) a SymR.[1] Navzdory podobnosti s nadrodinou AbrB byla rodina SymE nalezena výhradně v proteobakteriích.[1]
Vztah mezi SymE a SymR
The SymR antisense RNA je přepsal 3 nt za SymE spustit kodon což je důvod, proč je promotor SymR považován za vložený do SymE kodon.[2] Ve výsledku blokuje SymR RNA překlad SymE vazbou antisense, což naznačuje, že to nakonec vede k degradaci mRNA SymR.[4] Aminokyselina analýza dospěla k závěru, že se z SymE může vyvinout protein štěpící RNA, který kvůli tomu vykazuje toxinové chování transkripční faktory nebo antitoxiny.[2] Na rozdíl od jiných běžných toxin-antitoxinových systémů je antitoxin SymR stabilnější než toxin SymE.[1]
Následující Poškození DNA, SOS odpověď potlačuje transkripce SymR RNA, což umožňuje toxinu SymE degradovat potenciálně poškozenou mRNA, dokud nebyla DNA opraveno.[1] Naopak, SymE je přísně potlačen LexA vazebná místa represoru, SymR a Lon proteáza.[2] Tyto tři faktory jsou přítomné na několika úrovních, kde se LexA podílí na transkripci downregulace „SymR RNA se účastní translace downregulace a Lon proteáza se podílí na proteinu degradace.[1][2] Rozsah represe na SymE závisí na aditivní síle LexA, SymR a Lon proteázy.[2] Syntéza SymE je celkově pomalá, protože její aktivita vysoce závisí na opravných proteinech DNA.[2] V buněčném prostředí mitomycin C. poškozuje DNA, což vede k nadměrné expresi mRNA SymE k zahájení opravy DNA.[5]
Toxicita
The nadměrná exprese SymE prokázalo negativní účinky na růst buňky tvořící kolonie při testování in vitro.[1] SymE vystavuje své toxicita potlačením globálního překladu v rámci buňka, štěpení mRNA podobným způsobem jako MazF další toxin.[6] Kvantitativní Northern blot experimenty ukázaly, že SymR RNA je v buňkách přítomna při 10násobku koncentrace mENA SymE (0,02 fmol μg−1 a 0,2 fmol μg−1).[1]
Struktura
SymE
Toxin SymE se skládá ze 113 aminokyseliny.[5] Při hodnocení aminokyselinová sekvence a terciární struktura ze SymE byly nalezeny silné podobnosti, které se podobají AbrB nadčeleď.[1] Tato nadčeleď funguje hlavně jako transkripční faktory nebo antitoxiny; podobnost SymE s primární sekvence a terciární struktura nadrodiny AbrB naznačuje, že proteiny SymE zažily evoluční posun z a transkripční faktor nebo antitoxin na protein asociovaný s RNA, který vykazuje toxinové chování.[1] Mezi proteinovou strukturou nadrodiny AbrB a proteinovou strukturou SymE existuje několik klíčových hydrofobní zbytky které jsou v -spirála ve středu bílkoviny i pramen -1.[1] Přes tyto klíčové podobnosti vykazuje SymE polární zbytky, které se nenacházejí v obecné struktuře nadrodiny AbrB, což naznačuje, že tyto zbytky mohou hrát roli ve schopnosti štěpení RNA SymE.[1]


ŠVÝCARSKÝ MODEL obsahuje více než několik experimentálních struktur a teoretických homologie modely, které definují určité aspekty SymE primární sekvence a terciární struktura. The UniProtKB přístupové číslo P39394 označuje obecnou strukturu toxinu SymE v Escherichia coli (kmen K12).[1][7] V teoretickém modelu SWISS-MODEL SymE je -spirála obsahuje aminokyseliny G 44, Q 45, Ž 46, L 47, E 48, A 49 a A 50.[8][9][10][11][12] The pramen -1 obsahuje aminokyseliny G 55, T 56, A 57, PROTI 58, D 59, PROTI 60, K. 61, PROTI 62, Já 67, PROTI 68, L 69, T 70, A 71, Q 72, P 73 a P 74 s -otáčet se obsahující M 63, E 64, G 65 a C 66.[8][9][10][11][12]
SymR
SymR je antisense RNA což znamená jeho sekundární struktura má charakteristiku kmen a smyčka prvky i nepárové oblasti lemující strukturu.[13] Předpovězeno sekundární struktura SymR představuje smyčku obsahující nukleotidovou sekvenci CCAG.[4] Tato charakteristická smyčka je sdílena s proteiny lstR-1 a OhsC RNA a předpokládá se, že je a vazebné místo pro jiné proteiny.[4] V současné době nejsou na serveru žádné známé soubory RCSB proteinová banka nebo Úložiště SWISS-MODEL které naznačují předpokládanou terciární strukturu SymR.
Viz také
Reference
- ^ A b C d E F G h i j k l m n Kawano M, Aravind L, Storz G (květen 2007). „Antisense RNA řídí syntézu toxinu vyvolaného SOS, který se vyvinul z antitoxinu“. Molekulární mikrobiologie. 64 (3): 738–54. doi:10.1111 / j.1365-2958.2007.05688.x. PMC 1891008. PMID 17462020.
- ^ A b C d E F G Kawano M (prosinec 2012). „Odlišně se překrývající cis-kódovaná antisense RNA regulující toxin-antitoxinové systémy z E. coli: hok / sok, ldr / rdl, symE / symR“. RNA Biology. 9 (12): 1520–7. doi:10,4161 / rna.22757. PMID 23131729.
- ^ Kawano M, Reynolds AA, Miranda-Rios J, Storz G (2005). "Detekce 5'- a 3'-UTR odvozených malých RNA a cis-kódovaných antisense RNA v Escherichia coli". Výzkum nukleových kyselin. 33 (3): 1040–50. doi:10.1093 / nar / gki256. PMC 549416. PMID 15718303.
- ^ A b C Fozo EM, Hemm MR, Storz G (prosinec 2008). „Malé toxické proteiny a antisense RNA, které je potlačují“. Recenze mikrobiologie a molekulární biologie. 72 (4): 579–89, obsah. doi:10.1128 / MMBR.00025-08. PMC 2593563. PMID 19052321.
- ^ A b Brielle R, Pinel-Marie ML, Felden B (duben 2016). „Spojení bakteriálních toxinů typu I s jejich účinky“ (PDF). Současný názor v mikrobiologii. Regulace buněk. 30: 114–121. doi:10.1016 / j.mib.2016.01.009. PMID 26874964.
- ^ Gerdes K, Wagner EG (duben 2007). "RNA antitoxiny". Současný názor v mikrobiologii. 10 (2): 117–24. doi:10.1016 / j.mib.2007.03.003. PMID 17376733.
- ^ Konsorcium UniProt (2020). „UniProtKB - P39394 (SYME_ECOLI)“. uniprot.org. Archivováno z původního dne 10. července 2007. Citováno 4. května 2020.
- ^ A b Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R a kol. (Červenec 2018). „SWISS-MODEL: homologické modelování proteinových struktur a komplexů“. Výzkum nukleových kyselin. 46 (W1): W296 – W303. doi:10.1093 / nar / gky427. PMC 6030848. PMID 29788355.
- ^ A b Guex N, Peitsch MC, Schwede T (červen 2009). „Automatizované srovnávací modelování proteinové struktury pomocí SWISS-MODEL a Swiss-PdbViewer: historická perspektiva“. Elektroforéza. 30 Suppl 1 (S1): S162-73. doi:10.1002 / elps.200900140. PMID 19517507.
- ^ A b Bienert S, Waterhouse A, de Beer TA, Tauriello G, Studer G, Bordoli L, Schwede T (leden 2017). „Úložiště SWISS-MODEL - nové funkce a funkce“. Výzkum nukleových kyselin. 45 (D1): D313 – D319. doi:10.1093 / nar / gkw1132. PMC 5210589. PMID 27899672.
- ^ A b Studer G, Rempfer C, Waterhouse AM, Gumienny R, Haas J, Schwede T (duben 2020). „Omezení QMEANDisCo-vzdálenost aplikovaná na odhad kvality modelu“. Bioinformatika. 36 (8): 2647. doi:10.1093 / bioinformatika / btaa058. PMC 7178391. PMID 32048708.
- ^ A b Bertoni M, Kiefer F, Biasini M, Bordoli L, Schwede T (září 2017). "Modelování proteinové kvartérní struktury homo- a hetero-oligomerů nad binární interakce pomocí homologie". Vědecké zprávy. 7 (1): 10480. Bibcode:2017NatSR ... 710480B. doi:10.1038 / s41598-017-09654-8. PMC 5585393. PMID 28874689.
- ^ Brenner SX, Miller JH. Encyklopedie genetiky. San Diego. ISBN 0-12-227080-0. OCLC 48655705.
Další čtení
- Buts L, Lah J, Dao-Thi MH, Wyns L, Loris R (prosinec 2005). „Moduly toxin-antitoxin jako správce bakteriálního metabolického stresu“. Trendy v biochemických vědách. 30 (12): 672–9. doi:10.1016 / j.tibs.2005.10.004. PMID 16257530.
- Gerdes K, Christensen SK, Løbner-Olesen A (květen 2005). "Prokaryotický toxin-antitoxinový stresový lokus". Recenze přírody. Mikrobiologie. 3 (5): 371–82. doi:10.1038 / nrmicro1147. PMID 15864262.
- Lewis LK, Harlow GR, Gregg-Jolly LA, Mount DW (srpen 1994). "Identifikace vysoce afinitních vazebných míst pro LexA, která definují nové geny indukovatelné poškození DNA v Escherichia coli". Journal of Molecular Biology. 241 (4): 507–23. doi:10.1006 / jmbi.1994.1528. PMID 8057377.
- Christensen SK, Pedersen K, Hansen FG, Gerdes K (září 2003). „Toxin-antitoxinové lokusy jako prvky reakce na stres: ChpAK / MazF a ChpBK štěpí přeložené RNA a působí proti nim tmRNA.“ Journal of Molecular Biology. 332 (4): 809–19. doi:10.1016 / S0022-2836 (03) 00922-7. PMID 12972253.
- Engelberg-Kulka H, Glaser G (1999). "Moduly závislosti a programovaná buněčná smrt a antideath v bakteriálních kulturách". Výroční přehled mikrobiologie. 53: 43–70. doi:10.1146 / annurev.micro.53.1.43. PMID 10547685.
- Cherepanov PP, Wackernagel W (květen 1995). „Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-katalyzated excision of the antibiotic-rezistence determinant“. Gen. 158 (1): 9–14. doi:10.1016 / 0378-1119 (95) 00193-A. PMID 7789817.
- Anantharaman V, Aravind L (2003). „Nová spojení v síti prokaryotických toxinů a antitoxinů: vztah s eukaryotickým nesmyslem zprostředkovaným systémem rozpadu RNA“. Genome Biology. 4 (12): R81. doi:10.1186 / gb-2003-4-12-r81. PMC 329420. PMID 14659018.