Folikulární B pomocné T buňky - Follicular B helper T cells
Folikulární B pomocné T buňky (také známý jako spravedlivý folikulární pomocné T buňky nebo TFH), jsou antigen -zkušený CD4+ T buňky nachází se na periferii uvnitř B buňka folikuly sekundárních lymfoidních orgánů, jako je lymfatické uzliny, sleziny a Peyerovy náplasti, a jsou identifikovány podle jejich konstitutivního vyjádření B buňka receptor pro navádění folikulů CXCR5.[1] Po buněčné interakci a křížové signalizaci s jejich příbuznými folikulární (Fo B) B buňky, TFH buňky spouští vznik a údržbu zárodečná centra prostřednictvím výrazu Ligand CD40 (CD40L) a sekrece IL-21[2] a IL-4.[3] TFH buňky také migrují do těchto semen zárodečná centra, převážně složené z rychle se dělících a mutovat B buňky. V zárodečných centrech TFH buňky hrají rozhodující roli při zprostředkování výběru a přežití B buněk, které dále diferencují buď na speciální plazmatické buňky schopné produkovat vysoce afinitní protilátky proti cizímu antigenu, nebo paměťové B buňky schopný rychlé imunitní reaktivace v budoucnu, pokud dojde k opětovnému výskytu stejného antigenu. TFH také se předpokládá, že buňky usnadňují negativní selekci potenciálně autoimunitní - způsobující mutované B buňky v zárodečné centrum. Avšak biomechanismy, kterými TFH buňky zprostředkovávají zárodečné centrum tolerance dosud plně pochopeny.
Je možné, že TFH buňky mohou vznikat jako větve v diferenciačních drahách Th1 a Th2, ale jejich přesný vztah linií k dalšímu efektorovému CD4+ Podskupiny T buněk jsou stále nejisté. Nedávné studie však ukázaly, že TFH mají odlišné profily genové exprese, podporující teorii, že TFH jsou podmnožinou CD4+ T buňky odlišné od Čt-1, Th-2, Čt-17 nebo Tregs.[4][5]
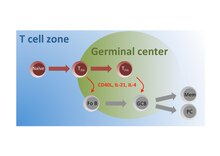
Podskupina naivních T buněk v zóně T buněk je aktivována antigenem a migrují do folikulů, kde se diferencují na TFH buňky, které interagují s folikulárními B (Fo B) buňkami a instruují je, aby podstoupily izotypovou změnu, somatickou hypermutaci a rychlé buněčné dělení na zárodečná centra semen (GC). V těchto zárodečných centrech TFH buňky nadále poskytují pomoc GC B buňkám, aby usnadnily jejich produkci vysoce afinitních protilátek produkujících plazmatických buněk (PC) a B buněk s dlouhou životností (Mem).
Biomolekulární charakterizace
The indukovatelný kostimulátor T-buněk (CD278 nebo ICOS) je prokázáno, že poskytuje zvláště kritický signál pro TFH buňky, protože experimentální myši s deficitem ICOS nejsou schopné vyvinout žádný TFH.[6] Dále bylo prokázáno, že ICOS indukuje sekreci IL-21 cytokin aktivovaným CD4+ T buňky a že IL-21 hraje klíčovou roli ve vývoji TFH buňky a zárodečná centra.[7][8] Taky Bcl-6 je transkripční faktor identifikovaný v TFH buňky, ale může mít role, které přesahují tuto podmnožinu, protože to bylo také zapleteno do paměti CD8+ Vývoj T buněk.[9]
V zárodečných centrech se zkušenostmi s antigeny TFH buňky rychle upregulují expresi CD40L, která váže a stimuluje povrchový receptor B buněk CD40.[10] TFH závislé na buňkách parakrin aktivace B-buněk CD40 vede k přežití a diferenciaci B-buněk, včetně indukce AID (aktivací (cytidin) deamináza ).[11] AID výraz (kódovaný pomocí AICDA Gen) způsobí, že protilátky B-lymfocytů přepnou z IgM / IgD na jiné izotypy a disky protilátek somatická hypermutace během klonální proliferace. Změněné protilátky získávají lepší efektorové funkce a hypermutovaná protilátka vykazuje vyšší afinitu k antigenu.
Třídy TFH buňky
TFH buňky vytvořené brzy v rodících se stadiích a zárodečné centrum reakce se formálně nazývají pre-TFH buňky. Jsou jedinečně nalezeny převážně na hranici zóny T buněk, která splývá s folikuly B buněk a zárodečnými centry. Pre-TFH buňky jsou funkčně velmi podobné ostatním TFH buňky při usnadňování zárodečných center B buněčných reakcí, ale navíc jsou také schopné řídit vývoj folikulárních B buněk v sousedství zárodečných center a mimo ně, aby produkovaly rychle reagující, ale netrvalé plazmatická buňka řízené protilátkové odpovědi (známé jako extrafolikulární odpověď).
Ty TFH buňky specificky umístěné ve zralém germinálním centru jsou někdy označovány jako GC TFH buňky (pro Germinal Center TFH buňky), aby se odlišily od pre-TFH buňky.[12][13] V této populaci GC Tfh buněk je také menší podtřída, která exprimuje gen Foxp3, kódující transkripční faktor. Tato malá diskrétní subpopulace buněk, nazývaná TFR buňky (pro T folikulární regulační buňky), jsou důležité při kontrole a omezování rozsahu normálních reakcí germinálních center tak, aby se zabránilo potenciálu produkovat abnormálně mutované nebo samovolně autoimunitní -sociální protilátky.[14] Proto TFR buňky jsou jedinečným inhibičním vlivem během reakce germinálního centra.
Zatímco TFH buňky se nacházejí primárně v sekundárních lymfoidních orgánech, malá část cirkuluje v krvi a jsou označovány jako „periferní“ T folikulární pomocné buňky (pTFH). Tyto buňky lze identifikovat podle jejich exprese IL-21 po stimulaci.[15]
Lékařský význam
Generování trvalé imunitní paměti
TFH Buňky jsou považovány za nepostradatelnou podmnožinu T buněk při generování a údržbě zárodečné centrum odpovědi. Proto v nepřítomnosti TFH buňky, podobné aktivaci B buněk antigeny nezávislými na T-buňkách, se vytvoří rychlý výbuch produkce nízkoafinitních plazmatických buněk, ale to nevede k indukci germinálního centra ani k povolení protilátky afinitní zrání nebo rozlišení efektivní paměťové B buňky které jsou nezbytné pro posílení těla proti následným infekcím. Specificky jsou paměťové B buňky závislé na zárodečných centrech hnacím motorem vyvolání produkce protilátek během sekundární imunitní odpovědi. Proto je správná aktivace a vývoj TFH buněk je ústřední pro účinnost očkování a vakcína design pro indukci dlouhodobého imunita. V bangladéšské populační studii pacientů infikovaných Vibrio cholera a zdravých lidských dobrovolníků, kterým byla podána stávající vakcína proti choleře,[16] vzpomínka TFH reakce specificky proti antigenu cholery korelovala s další produkcí protilátek B buňkami.
S normálním stárnutím dochází k postupnému snižování imunitního systému těla. Tento jev se nazývá imunosenescence je do značné míry způsoben poklesem funkce T buněk, včetně kapacity pro TFH buňky správně podporovat zárodečné centrum odpovědi.[17] To může být částečně způsobeno nižší CD40L hladiny na povrchu buňky TFH buňky ve věku.[18]
Vyhněte se autoimunitě
Nezaškrtnuté nebo nadměrně aktivní TFH buněčné imunitní odpovědi mají potenciál připojit neoprávněná zárodečná centra, složená z aberantně mutovaných B buňky které mohou být zprostředkovány protilátkami autoimunitní onemocnění. Zvýšené hladiny TFHBuňky podobné buňkám lze detekovat v krvi podskupiny lidských pacientů s systémový lupus erythematodes (SLE) a Sjögrenův syndrom.[19] Vědecké důkazy naznačující TFH buňky mohou definitivně způsobit autoimunitu u lidí zůstává neúplné.
Reference
- ^ Fazilleau; Mark, L; McHeyzer-Williams, LJ; McHeyzer-Williams, MG; et al. (Březen 2009). "Folikulární pomocné T buňky: linie a umístění". Imunita. 30 (3): 324–35. doi:10.1016 / j.immuni.2009.03.003. PMC 2731675. PMID 19303387.
- ^ Seo; Youn, J; Kim, PH; et al. (Květen 2009). „IL-21 zajišťuje expresi izotypu IgA indukovanou TGF-beta 1 v myších Peyerových náplastech“. J. Leukoc Biol. 85 (5): 744–50. doi:10.1189 / jlb.0708450. PMID 19168593.
- ^ Zaretsky; Taylor, JJ; King, IL; Marshall, FA; Mohrs, M; Pearce, EJ; et al. (Květen 2009). "T folikulární pomocné buňky se odlišují od Th2 buněk v reakci na helminthové antigeny". J Exp Med. 206 (5): 991–9. doi:10.1084 / jem.20090303. PMC 2715032. PMID 19380637.
- ^ Chtanova; Tangye, SG; Newton, R; Frank, N; Hodge, MR; Rolph, MS; MacKay, ČR; et al. (Červenec 2004). „Pomocné T folikulární buňky exprimují výrazný transkripční profil, což odráží jejich roli jako efektorových buněk jiných než Th1 / Th2, které pomáhají B buňkám.“. J. Immunol. 173 (1): 68–78. doi:10,4049 / jimmunol.173.1.68. PMID 15210760.
- ^ Nurieva RI, Chung Y, Hwang D, Yang XO, Kang HS, Ma L, Wang YH, Watowich SS a kol. (2008). „Generování T folikulárních pomocných buněk je zprostředkováno interleukinem-21, ale nezávisle na T-pomocných liniích 1, 2 nebo 17 buněk“. Imunita. 29 (1): 138–49. doi:10.1016 / j.immuni.2008.05.009. PMC 2556461. PMID 18599325.
- ^ Akiba; Takeda, K; Kojima, Y; Usui, Y; Harada, N; Yamazaki, T; Ma, J; Tezuka, K; et al. (Srpen 2005). „Role ICOS při udržování folikulárních B pomocných T-buněk CXCR5 + in vivo“. J. Immunol. 175 (4): 2340–8. doi:10,4049 / jimmunol.175.4.2340. PMID 16081804.
- ^ Bauquet; Jin, H; Paterson, AM; Mitsdoerffer, M; Ho, IC; Sharpe, AH; Kuchroo, VK (únor 2009). „Kostimulační molekula ICOS reguluje expresi c-Maf a IL-21 ve vývoji folikulárních T pomocných buněk a buněk TH-17“. Nat Immunol. 10 (2): 167–75. doi:10.1038 / ni.1690. PMC 2742982. PMID 19098919.
- ^ Vogelzang; McGuire, HM; Yu, D; Sprent, J; MacKay, ČR; King, C (červenec 2008). „Základní role interleukinu-21 při tvorbě pomocných T folikulárních buněk“. Imunita. 29 (1): 127–37. doi:10.1016 / j.immuni.2008.06.001. PMID 18602282.
- ^ Ichii; Sakamoto, A; Hatano, M; Okada, S; Toyama, H; Taki, S; Arima, M; Kuroda, Y; Tokuhisa, T; et al. (Červen 2002). "Role pro Bcl-6 při generování a údržbě paměťových CD8 + T buněk". Nat Immunol. 3 (6): 558–63. doi:10.1038 / ni802. PMID 12021781.
- ^ Kim, JR; Lim, HW; Kang, SG; Hillsamer, P; Kim, CH (4. února 2005). „Lidské zárodečné centrum-T buňky CD57 + jsou hlavními pomocníky pro buňky GC-B a indukují rekombinaci přepínání tříd“. Imunologie BMC. 6: 3. doi:10.1186/1471-2172-6-3. PMC 548684. PMID 15694005.
- ^ McHeyzer-Williams, M; Okitsu, S; Wang, N; McHeyzer-Williams, L (9. prosince 2011). "Molekulární programování paměti B buněk". Recenze přírody. Imunologie. 12 (1): 24–34. doi:10.1038 / nri3128. PMC 3947622. PMID 22158414.
- ^ Yusuf I, Kageyama R, Monticelli L, Johnston RJ, Ditoro D, Hansen K, Barnett B, Crotty S (červenec 2010). „Produkce IL-4 germinálního centra T folikulární pomocné buňky závisí na signalizaci receptoru molekuly lymfocytární aktivace (CD150)“. J. Immunol. 185 (1): 190–202. doi:10,4049 / jimmunol.0903505. PMC 2913439. PMID 20525889.
- ^ Ramiscal RR, Vinuesa CG (březen 2013). "Podskupiny T-buněk v zárodečném centru". Immunol. Rev. 252 (1): 146–55. doi:10.1111 / imr.12031. PMID 23405902.
- ^ Wollenberg I, Agua-Doce A, Hernández A, Almeida C, Oliveira VG, Faro J, Graca L (listopad 2011). "Regulace reakce zárodečného centra pomocí Foxp3 + folikulárních regulačních T buněk". J. Immunol. 187 (9): 4553–60. doi:10,4049 / jimmunol. 1101328. PMID 21984700.
- ^ Schultz, Bruce T .; Teigler, Jeffrey E .; Pissani, Franco; Oster, Alexander F .; Kranias, Gregory; Alter, Galit; Marovich, Mary; Eller, Michael A .; Dittmer, Ulf (2016-01-19). „Cirkulující HIV specifické buňky interleukinu-21 (+) CD4 (+) T představují periferní Tfh buňky s pomocnými funkcemi závislými na antigenu“. Imunita. 44 (1): 167–178. doi:10.1016 / j.immuni.2015.12.011. ISSN 1097-4180. PMID 26795249.
- ^ Arifuzzaman M, Rashu R, Leung DT, Hosen MI, Bhuiyan TR, Bhuiyan MS, Rahman MA, Khanam F, Saha A, Charles RC, LaRocque RC, Weil AA, Clements JD, Holmes RK, Calderwood SB, Harris JB, Ryan ET , Qadri F (srpen 2012). „Antigen-specifické paměťové T-buňky po očkování orálně zabitou cholerovou vakcínou u bengálských dětí a srovnání s odpověďmi u pacientů s přirozeně získanou cholerou“. Klinická a vakcínová imunologie. 19 (8): 1304–11. doi:10.1128 / CVI.00196-12. PMC 3416086. PMID 22739692.
- ^ Lefebvre JS, Maue AC, Eaton SM, Lanthier PA, Tighe M, Haynes L (2012). „Zestárlé mikroprostředí přispívá k funkčním vadám CD4 T buněk u myší související s věkem“. Stárnoucí buňka. 11 (5): 732–40. doi:10.1111 / j.1474-9726.2012.00836.x. PMC 3444657. PMID 22607653.
- ^ Perkey E, Miller RA, Garcia GG (2012). „Ex vivo enzymatické ošetření starých CD4 T buněk obnovuje funkci pomocných T buněk a zvyšuje produkci protilátek u myší“. J. Immunol. 189 (12): 5582–9. doi:10,4049 / jimmunol.1200487. PMC 3518580. PMID 23136198.
- ^ Řemeslo JE. (2012). "Folikulární pomocné T buňky v imunitě a systémové autoimunitě". Nat Rev Rheumatol. 8 (6): 337–47. doi:10.1038 / nrrheum.2012.58. PMC 3604997. PMID 22549246.