Cartorhynchus - Cartorhynchus
| Cartorhynchus | |
|---|---|
 | |
| 3D rekonstrukce lebky, při pohledu zleva dole, odhalující zuby a vnitřní střechu lebky | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Třída: | Reptilia |
| Clade: | †Ichthyosauromorpha |
| Clade: | †Ichthyosauriformes |
| Clade: | †Nasorostra |
| Rod: | †Cartorhynchus Motani et al., 2014 |
| Druh: | †C. lenticarpus |
| Binomické jméno | |
| †Cartorhynchus lenticarpus Motani et al., 2014 | |
Cartorhynchus (ve smyslu „zkrácený čenich“) je vyhynulý rod z brzy ichtyosauriform mořský plaz který žil během Raný trias epocha, asi před 248 miliony let. Rod obsahuje jediný druh, Cartorhynchus lenticarpus, pojmenovaný v roce 2014 Ryosuke Motani a kolegy z jedné téměř úplné kostry nalezené poblíž Chaohu, Provincie Anhui, Čína. Spolu s jeho blízkým příbuzným Sclerocormus, Cartorhynchus byla součástí diverzifikace mořských plazů, ke které došlo náhle (více než milion let) během Spathian podstavec, brzy po ničivém Událost vyhynutí permu a triasu, ale následně byli vyhnáni k zániku vulkanismem a změnami hladiny moře ze strany Střední trias.
Měřící asi 40 centimetrů (16 palců) dlouhý, Cartorhynchus bylo malé zvíře s ještěrkovitým tělem a krátkým trupem; pravděpodobně plaval úhořem při nízkých rychlostech. Jeho končetiny nesly rozsáhlou chrupavku a mohly se ohýbat jako ploutve, což jí umožňovalo chodit po zemi. Nejvýraznější rysy Cartorhynchus byly jeho krátký, zúžený čenich a jeho několik řad molární -jako zuby, které rostly na vnitřním povrchu čelistních kostí. Tyto zuby nebyly objeveny, dokud nebyl vzorek podroben CT skenování. Cartorhynchus pravděpodobně lovil bezobratlých s tvrdým krunýřem sání krmení, i když zatím není známo, jak přesně používal zuby směřující dovnitř. Bylo to jedno z až pěti nezávislých akvizic molárních zubů mezi ichtyosauriformami.
Objev a pojmenování

V roce 2011 jediný známý exemplář Cartorhynchus byl objeven v posteli 633 z druhé úrovně lomu Majiashan poblíž centra města Chaohu, Provincie Anhui, Čína; horninové vrstvy v tomto lomu patří hornímu členu Formace Nanlinghu.[1] Vzorek se skládá z téměř úplné kostry, kde chybí pouze část ocasu[2] a některé kosti z levé části zadní lebky. Konzervace vzorku pravděpodobně vyplynula z toho, že byl uložen v sedimentu pravou stranou dolů, takže levá strana byla vystavena prvkům. To přijalo číslo pole MT-II, a později číslo vzorku AGB 6257 na Geologické muzeum Anhui.[3]
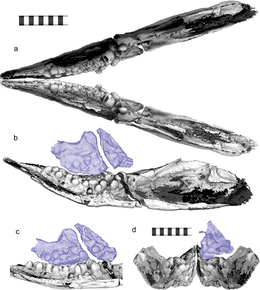
V roce 2014 byl vzorek popsán Ryosuke Motani a kolegy v Příroda jako zástupce nového rodu a druhu, Cartorhynchus lenticarpus. Odvodili druhové jméno Cartorhynchus z řeckých slov Kartos (καρτοσ, „zkráceno“) a rinchios (ρηψνχηοσ, "čenich") a konkrétní název lenticarpus z latinských slov lentus („flexibilní“) a zápěstí ("zápěstí"). Obě jména odkazují na anatomické vlastnosti, které by měl v životě.[1] Vzorek byl považován za bezzubý, dokud nebyl objeven izolovaný zub během dalších pokusů o odstranění horniny mezi uzavřenými čelistmi. Vzhledem k tomu, že vzorek byl příliš křehký, aby odhalil vnitřek čelistí, Jian-Dong Huang, Motani a další kolegové následně naskenovali a vykreslili vzorek ve 3D pomocí mikropočítačová tomografie (mikro-CT), provedeno ve společnosti Yinghua Testing Company v Šanghaj, Čína. V roce 2020 byly výsledky jejich následné práce zveřejněny v Vědecké zprávy.[3]
Popis

V době svého objevu Cartorhynchus byl nejmenší známý člen Ichthyosauriformes. Zachovaný exemplář měl délku 21,4 cm (8 palců); za předpokladu, že měl proporce ocasu srovnatelné s blízkými příbuznými, odhadovali Motani a kolegové délku celého těla 40 centimetrů (1 ft 4 v) a hmotnost 2 kilogramů (4,4 lb).[1][2]
Lebka
Cartorhynchus měl neobvykle krátký a zúžený čenich, který zabíral jen polovinu délky lebky, a hlubokou čelist. Špička čenichu byla široká pouze 6 milimetrů (0,2 palce).[3] Na rozdíl od většiny plazů jeho nosní kost dosáhl přední části čenichu. Díky tomu je také protáhlý premaxilla, jeho kostnaté nosní dírky byly umístěny relativně daleko vzadu na lebce, a jeho čelní kost také postrádal expanzi v zadním vnějším rohu. Všechny tyto charakteristiky byly sdíleny s jeho blízkým příbuzným Sclerocormus. Na rozdíl od toho však čelní kost nepřispívala do očního důlku Cartorhynchus; the prefrontální a postfrontální kosti se nesetkaly nad očním důlkem;[4] a umístění velkého otvoru pro epifýza na střeše lebky se lišila: byla v kontaktu mezi čelní a temenní kosti v Sclerocormus, ale pouze na parietals v Cartorhynchus. Cartorhynchus měl také charakteristicky velký hyoidní kost.[1][5]

Zpočátku to odvodili Motani a kolegové Cartorhynchus byl bezzubý; mikro-CT skenování však následně odhalilo přítomnost zaoblených, molární - jako zuby, které vyčnívají dovnitř téměř kolmo k dlouhé ose čelisti, a proto je zvenčí neviditelné. Všechny zuby byly buď úplně zploštělé nebo slabě špičaté a mnoho zubů mělo zúžení mezi kořenem a temenem. Na rozdíl od jiných ichthyosauriformů s molariformními zuby byly všechny korunky zubů „oteklé“ v podobném rozsahu. Na horní čelist (horní čelist) a zubař (dolní čelist), zuby byly uspořádány do tří řad, přičemž nejvzdálenější řada měla nejvíce a největší zuby; čelisti měly každý sedm, pět a pravděpodobně jeden zub, zatímco zubary měly každý deset, sedm a čtyři zuby. Pouze mezi ichtyosauriformami Cartorhynchus a Xinminosaurus mít více řad zubů.[6] Uspořádání zubů znamenalo, že přední přední spodní zuby by neměly žádný odpovídající horní zub, a také to, že dva dentary tvořící dolní čelist nemohly být pevně spojeny. Tuto vlastnost by sdíleli její blízcí příbuzní, bezzubí hupehsuchians.[3][7]
Postkraniální kostra

Cartorhynchus Zdá se, že měl 5 krční obratle a 26 zadní obratle, celkem 31 předsakrálních obratlů (obratel před jeho křížová kost nebo kyčle). Spolu s Sclerocormus (s 34 předsakrálními) a Chaohusaurus (s 36 předsakrálními), Cartorhynchus spadá do typického rozmezí pro suchozemská zvířata, na rozdíl od 40 až 80 předsakrálních běžných mezi více odvozený (specializované) ichtyopterygians.[1] Na rozdíl od Sclerocormus, neurální trny vyčnívající z horní části obratlů dovnitř Cartorhynchus byly relativně úzké a nakloněné místo široké a přírubové. Cartorhynchus lze také odlišit parafofýzami, vertebrálními procesy, které se kloubově spojují s žebry; jejich přední okraje splývaly s okraji obratlů.[5]
Oba Cartorhynchus a Sclerocormus měl těžce stavěné žebra, které byly nejhlubší u ramen, se širokými, zploštělými a silnostěnný žebra, jak je běžně vidět u raných členů linií druhotně vodních plazů.[1][8] Ichthyopterygia ztratila tato zploštělá žebra s výjimkou Mollesaurus.[9] Na spodní straně hrudníku je gastralia z Cartorhynchus byly tenké a podobné tyči, na rozdíl od zploštělého „koše“ Sclerocormus, ale oběma chyběla další dvojice symetrických prvků ve středové linii těla.[5]
Končetiny Cartorhynchus byly špatně zkostnatělý (pouze tři číslice ruky byly zkostnatělé) se široce rozmístěnými kostmi, zejména mezi kosti zápěstí (carpals) a číslice, což naznačuje přítomnost rozsáhlé chrupavky v končetinách. To by končetinám připomínalo ploutev. Přední končetiny ploutve Cartorhynchus byly zakřiveny dozadu, přičemž číslice byly nakloněny o 50 ° vzhledem k ose dlouhých kostí (zeugopodium), zatímco zadní končetiny byly zakřiveny dopředu. The stehenní kost z Cartorhynchus byl rovný a neroztažený na spodním konci. Sclerocormus měli podobné končetiny, až na to, že byly lépe osifikovány a jejich zachované zakřivení možná nebylo přirozené.[1][5]
Klasifikace

Nedostatek úplných fosilních pozůstatků má za následek nejasnost ohledně původu Ichthyopterygie, včetně ichtyosaury. Po mnoho let se předpokládalo, že se jejich fosilie náhle objevily v Střední trias se silnými vodními adaptacemi. Objev Cartorhynchus a Sclerocormus tuto mezeru částečně vyplnil. Fylogenetické analýzy provedené Motanim a jeho kolegy zjistili, že oba spolu úzce souvisí - tvořící a clade (skupina) volal Nasorostra - a na Ichthyopterygia, ke které nasorostrans tvořil sesterská skupina. Cartorhynchus a Sclerocormus spojily je jejich krátké čenichy, protáhlé nosní dírky, hluboké čelisti, čelní strany bez rozšíření, hrudní koše nejhlubší u ramen a lopatky (lopatky) širší na spodním konci než na horním konci.[1][5]
Začlenění nasorostranů do fylogenetických analýz také poskytlo důkazy na podporu hupehsuchianů jako blízkých příbuzných ichtyopterygiánů. V roce 2014 Motani a kolegové pojmenovali klad tvořený Nasorostrou a Ichthyopterygií jako Ichthyosauriformes a klade tvořený Ichthyosauriformes a Hupehsuchia jako Ichthyosauromorpha. Zejména jejich analýzy obnovily úzký vztah mezi těmito různými skupinami bez ohledu na to, zda byly z analýzy odstraněny charakteristiky spojené s vodními adaptacemi.[1][5] Tyto vlastnosti se mohly vyvinout homoplasiously (z konvergentní evoluce ) mezi více liniemi v důsledku podobných životních stylů, které mohou zkreslit fylogenetické analýzy a rekonstruovat je jako homologie (odvozeno od sdíleného původu). Přetrvávání těchto rekonstruovaných vztahů i po odstranění vodních charakteristik ukazuje na jejich robustnost.[10][11]
Níže je fylogenetický strom z fylogenetické analýzy publikované Huangem, Motanim a kolegy v roce 2019 v popisu Chaohusaurus brevifemoralis, je částečně reprodukován.
| Ichthyosauromorpha |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Evoluční historie
Vzhled ichthyosauromorfů byl součástí obnovy mořských ekosystémů po ničivých událostech Událost vyhynutí perm-trias. Obecně se věřilo, že mořské ekosystémy neobnovily svoji plnou rozmanitost až 5 až 10 milionů let po masovém vyhynutí a že mořští plazi se z vyhynutí zotavili pomaleji než jiné linie.[12][13] Objev mnoha rozmanitých faun mořských plazů vyskytujících se v raném triasu - včetně Cartorhynchus - následně ukázal, že tomu tak nebylo.[7][14] Zejména se zdá, že ichthyosauriforms se poprvé objevily během spathské podskupiny Olenekian etapa a rychle dosáhl vysokého bodu funkční rozmanitost v prvních milionech let jejich vývoje.[5] Obsadili různé výklenky navzdory relativně nízké druhové rozmanitosti,[15] včetně obou při dně (obydlí na dně) jako hupehsuchians a nasorostrans a pelagický (na volné vodě) druhy jako ichtyopterygians.[5]

Mnoho ichtyosauriformů z raného a Střední trias měl molariformní zuby (zobrazené ve fylogenetickém stromu výše), včetně Cartorhynchus; takové zuby naznačují stravu alespoň částečně založenou na tvrdých skořápkách.[16] V roce 2020 Huang a jeho kolegové provedli rekonstrukce stavu předků zubů mezi ichtyosauromorfy. Pravděpodobnostní metody naznačovaly, že zaoblené nebo ploché zuby se pravděpodobně vyvinuly samostatně pětkrát, zatímco metody založené na šetrnost navrhl, že se vyvinuli nezávisle třikrát až pětkrát. Huang a kolegové zjistili, že k vývoji molariformních zubů došlo u vodních živočichů mnohokrát nezávisle (včetně vícečetných linií monitorovat ještěrky, murény, a sparidní a cichlida ryby), a tedy frekvence mezi ichtyosauriformami není neobvykle vysoká. Pozorovali také, že rané triasové ichtyosauriformní bakterie měly obecně malé, zaoblené zuby; zuby středních triasových ichtyosauriformů byly rozmanitější co do velikosti a tvaru, což koreluje se zvýšenou rozmanitostí bezobratlých.[17] Navrhli tedy, že diverzifikace ichthyosauriformů byla částečně způsobena vývojem kořisti s tvrdou skořápkou.[3]
Hupehsuchians a nasorostrans však nakonec vyhynuli na hranici mezi raným a středním triasem a vytvořili „taxonomické úzké místo“. Na hranici se mění hladina moře[18] a vulkanismus[19] vedlo ke špatně okysličeným oceánům a vytvářelo charakteristiku podpis izotopu uhlíku z rozpadajícího se organického materiálu ve vrstvách hornin na hranici.[20] Ichthyosauriforms se po tomto obratu nezotavily v rozmanitosti,[21] s Sauropterygia a Saurosphargidae pohánět druhou vlnu diverzifikace trvající tři až pět milionů let.[5][15]
Paleobiologie
Strava
Motani a kolegové předpokládali v roce 2014, že Cartorhynchus byl sací podavač který se napájel soustředěním tlaku v úzkém čenichu. To bylo dále podpořeno robustností jeho hyoidního a hyobranchiálního prvku (který by zakotvil jazyk) a jejich nesprávným pozorováním bezzubosti.[1] Podobné závěry byly následně učiněny pro Sclerocormus.[5] Shastasaurus a Shonisaurus předtím byl interpretován jako jejichtyosauři krmení sáním,[22] ale kvantitativní analýza triasových a jurských ichtyosaurů, kterou provedli Motani a kolegové v roce 2013, ukázala, že žádný z nich neměl dostatečně robustní hyobranchiální kosti ani dostatečně úzké čenichy, aby umožnil sání.[23]
Objev molariformních zubů v Cartorhynchus vedl Huang a kolegy k tomu, aby v roce 2020 dospěli k závěru Cartorhynchus byl durophagous krmení kořistí s tvrdou skořápkou. Poznamenali, že to neodporuje životnímu stylu krmení; některé sparidní ryby jsou jak durofágní, tak sání.[24] Navrhli to však Cartorhynchus bylo by omezeno na krmení malou kořistí. Pokud jde o vodorovnou orientaci zubů, pozorovali opotřebitelné povrchy, které naznačovaly, že po stranách zubů uzavřený místo korun. Poznamenali však, že zuby jsou strukturálně nejsilnější na špičkách, ne na bocích, pod špičkou zdůrazňuje drcení kousnutí.[25] Navrhli, že během konzervace mohla být zkroucená čelist nebo měkké tkáně kolagen které v životě zvedly zuby, mohly být ztraceny, i když připustily, že ani jedna z hypotéz nevysvětlí vzorce opotřebení. Nakonec poznamenali, že dolní zuby bez odpovídajících horních zubů byly neobvyklé; nevykazují žádné vzorce opotřebení a neexistují žádné důkazy o svalových mechanismech, které by umožnily použití obou čelistí proti sobě. Proto vyvodili, že tyto zuby pravděpodobně nebyly použity proti jiným zubům.[3]
Končetiny a pohyb

Cartorhynchus měl špatně osifikované končetiny navzdory své dobře osifikované lebce a obratlům. Motani a kolegové však v roce 2014 navrhli, že to byl ještě dospělý člověk, protože mnoho časně odlišných členů linií mořských plazů má špatně zkostnatělé končetiny paedomorphosis (uchování nezralých znaků do dospělosti), i když zcela neodmítli možnost, že šlo o mladistvého kvůli existenci pouze jednoho exempláře.[1]
V případě Cartorhynchus„Motani a kolegové navrhli, aby mu velké přední končetiny připomínající ploutve umožnily pohyb po souši, čímž se staly obojživelnými. Rozsáhlá chrupavka v zápěstí by umožnila ohnout ploutev bez lokte; juvenilní mořské želvy mají podobně chrupavkovité ploutve, které používají k pohybu na souši.[26] Ačkoli jeho ploutev by nebyl zvlášť silný, Cartorhynchus byl relativně lehký, s poměrem tělesné hmotnosti k ploutvi menší než u Chaohusaurus. Zakřivené ploutve by jim umožňovaly držet je blízko u těla, čímž by se zvětšily jejich mechanická výhoda. Motani a kolegové navrhli, že další rysy Cartorhynchus také by pomohlo obojživelnému životnímu stylu, včetně krátkého kmene a čenichu a zesílených žeber (která by sloužila jako zátěž, stabilizace zvířete ve vodách na pobřeží).[1]

Studie Susany Gutarry a jejích kolegů z roku 2019 použila výpočetní simulace k odhadu energetických nákladů na plavání v ichtyosauriformách. Brzy se rozbíhající ichtyosauriformní s těly ještěrkovitých tvarů a protáhlými, bezchybnými ocasy Cartorhynchus, by byl zaměstnán anguilliformní (úhořovité) plavání, zatímco pozdější ichtyosauriformní s hlubšími těly podobnými rybám a dobře definovaným ocasním motolicím by se hodily carangiform plavání (jako makrela). Obecně se má za to, že plavání anguilliformem je méně účinné než plavání carangiform.[27][28][29] Gutarra a kolegové skutečně zjistili, že energetické náklady na plavání rychlostí 1 metr za sekundu (3,3 ft / s) byly 24 až 42krát vyšší v Cartorhynchus než ichtyosaurus Ophthalmosaurus (v závislosti na tom, zda je započítán plavecký režim) a jeho součinitel odporu vzduchu byl o 15% vyšší než v delfín skákavý. Nicméně, Cartorhynchus by pravděpodobně plaval pomalejšími rychlostmi vyžadujícími nižší efektivitu a výhody plavání v carangiform později byly větší ichtyosauriformní kompenzovány také zvětšením velikosti těla.[30]
Paleoekologie

Postel 633 v lomu Majiashan, lokalita kde Cartorhynchus byla nalezena, je vrstva šedé stříbrný (jíl nesoucí) vápenec nachází se 13 metrů nad základnou horního člena formace Nanlinghu. Je definován nahoře i dole vrstvami nažloutlého marls. Ve smyslu amonit biostratigrafie, tato postel patří Podkolumbiti pásmo.[1][31] Pro vrstvy olenekianského věku vystavené v lomu Majiashan byly vytvořeny odhady data s vysokým rozlišením na základě izotopových záznamů uhlík-13 cyklistika a spektrální gama paprsek protokoly (které měří množství záření ve skalách astronomického původu); Zejména postel 633 byla odhadnuta na 248,41 Ma ve věku.[32][33][34] Během středního triasu byly vrstvy Chaohu uloženy v oceánská pánev relativně daleko od pobřeží, které bylo na jihu ohraničeno mělčími vodami a karbonátové platformy, a na severu a kontinentální svah a hlubší povodí.[35][36]
Ichthyosauriforms Sclerocormus a Chaohusaurus se oba nacházejí v lomu Majiashan spolu s Cartorhynchus; Sclerocormus je známo z mladší postele 719 (248,16 Ma), zatímco Chaohusaurus se nachází v obou postelích.[32] Sauropterygian Majiashanosaurus je známo z postele 643.[37] Rozmanitost ryb v lomu Majiashan je horší než v jiných lokalitách; nejběžnější ryba je Chaohuperleidus, nejstarší známý člen Perleidiformes, ale druh širokého rozsahu Saurichthys a několik nepopsaných ryb je také neznámých.[38][39] Potenciální kořist bezobratlých pro Cartorhynchus zahrnují malé amonity a mlži a thylacocephalan členovec Ankitokazocaris.[3][31]
Viz také
Reference
- ^ A b C d E F G h i j k l m Motani, R .; Jiang, D.-Y .; Chen, G.-B .; Tintori, A .; Rieppel, O .; Ji, C .; Huang, J.-D. (2014). "Bazální ichtyosauriform s krátkým čenichem z dolního triasu v Číně". Příroda. 517 (7535): 485–488. Bibcode:2015 Natur.517..485M. doi:10.1038 / příroda13866. PMID 25383536. S2CID 4392798.
- ^ A b Perkins, Sid (2014). „Jak dostal jejich ploutev jejich ploutve“. Věda. Citováno 8. května 2020.
- ^ A b C d E F G Huang, J.-D .; Motani, R .; Jiang, D.-Y .; Ren, X.-X .; Tintori, A .; Rieppel, O .; Zhou, M .; Hu, Y.-C .; Zhang, R. (2020). „Opakovaný vývoj durofágie během radiace ichtyosaurů po masovém vyhynutí indikovaný skrytým chrupem“. Vědecké zprávy. 10 (1): 7798. Bibcode:2020NatSR..10.7798H. doi:10.1038 / s41598-020-64854-z. PMC 7210957. PMID 32385319.
- ^ Zhou, M .; Jiang, D.-Y .; Motani, R .; Tintori, A .; Ji, C .; Sun, Z.-Y .; Ni, P.-G .; Lu, H. (2017). „Lebeční osteologie odhalená trojrozměrně zachovanými lebkami raného triasu ichthyosauriform Chaohusaurus chaoxianensis (Reptilia: Ichthyosauromorpha) z Anhui v Číně “. Časopis paleontologie obratlovců. 37 (4): e1343831. doi:10.1080/02724634.2017.1343831. S2CID 90332288.
- ^ A b C d E F G h i j Jiang, D.-Y .; Motani, R .; Huang, J.-D .; Tintori, A .; Hu, Y.-C .; Rieppel, O .; Fraser, N.C .; Ji, C .; Kelley, N.P .; Fu, W.-L .; Zhang, R. (2016). „Velký aberantní stonek ichtyosauriform, který naznačuje časný vzestup a zánik ichtyosauromorfů v důsledku vyhynutí na konci permu“. Vědecké zprávy. 6: 26232. Bibcode:2016NatSR ... 626232J. doi:10.1038 / srep26232. PMC 4876504. PMID 27211319.
- ^ Jiang, D .; Motani, R .; Hao, W .; Schmitz, L .; Rieppel, O .; Sun, Y .; Sun, Z. (2008). „Nový primitivní ichtyosaurián (Reptilia, Diapsida) ze středního triasu z Panxian, Guizhou, jihozápadní Čína a jeho pozice v biotickém zotavení triasu“. Pokrok v přírodních vědách. 18 (10): 1315–1319. doi:10.1016 / j.pnsc.2008.01.039.
- ^ A b Motani, R .; Chen, X.H .; Jiang, D.-Y .; Cheng, L .; Tintori, A .; Rieppel, O. (2015). „Výpad krmení u raných mořských plazů a rychlý vývoj cechů na krmení mořských tetrapodů“. Vědecké zprávy. 5: 8900. Bibcode:2015NatSR ... 5E8900M. doi:10.1038 / srep08900. PMC 4354009. PMID 25754468.
- ^ Houssaye, A. (2009). ""Pachyostóza „u vodních plodů: recenze“. Integrativní zoologie. 4 (4): 325–340. doi:10.1111 / j.1749-4877.2009.00146.x. PMID 21392306.
- ^ Talevi, M .; Fernández, M.S. (2012). „Neočekávaná skeletální histologie ichtyosaura ze střední jury v Patagonii: důsledky pro vývoj mikrostruktury kostí mezi sekundárními vodními tetrapody“. Naturwissenschaften. 99 (3): 241–244. Bibcode:2012NW ..... 99..241T. doi:10.1007 / s00114-012-0886-4. PMID 22290413. S2CID 2223683.
- ^ Chen, X.-H .; Motani, R .; Cheng, L .; Jiang, D.-Y .; Rieppel, O. (2014). „Tajemný mořský plaz Nanchangosaurus z dolního triasu Hubei v Číně a z fylogenetických příbuzností Hupehsuchia “. PLOS ONE. 9 (7): e102361. Bibcode:2014PLoSO ... 9j2361C. doi:10.1371 / journal.pone.0102361. PMC 4094528. PMID 25014493.
- ^ Motani, R .; Jiang, D.-Y .; Tintori, A .; Rieppel, O .; Chen, G.-B .; Vy, H. (2015). „První důkazy o tom, že centralia v Ichthyopterygii opakuje zkreslení od paedomorfních postav na fylogenetické rekonstrukci mořských plazů“. Časopis paleontologie obratlovců. 35 (4): e948547. doi:10.1080/02724634.2014.948547. S2CID 83576278.
- ^ Chen, Z.Q .; Benton, M. J. (2012). „Načasování a schéma biotického zotavení po masovém vyhynutí na konci permu“. Nature Geoscience. 5 (6): 375–383. Bibcode:2012NatGe ... 5..375C. doi:10.1038 / ngeo1475.
- ^ Liu, J .; Hu, S.X .; Rieppel, O .; Jiang, D.-Y .; Benton, M. J.; Kelley, N.P .; Aitchison, J.C .; Zhou, C .; Wen, W .; Huang, J .; Xie, T .; Lv, T. (2014). „Gigantický nothosaur (Reptilia: Sauropterygia) ze středního triasu JZ Číny a jeho důsledky pro biotické zotavení triasu“. Vědecké zprávy. 4: 7142. Bibcode:2014NatSR ... 4E7142L. doi:10.1038 / srep07142. PMC 4245812. PMID 25429609.
- ^ Benton, M. J.; Zhang, Q .; Hu, S .; Chen, Z.-Q .; Wen, W .; Liu, J .; Huang, J .; Zhou, C .; Xie, T .; Tong, J .; Choo, B. (2013). „Výjimečné bioty obratlovců z čínského triasu a expanze mořských ekosystémů po hromadném vyhynutí permotriasu“. Recenze vědy o Zemi. 125: 199–243. Bibcode:2013ESRv..125..199B. doi:10.1016 / j.earscirev.2013.05.014.
- ^ A b Stubbs, T.L .; Benton, M.J. (2016). „Ekomorfologická diverzifikace druhohorních mořských plazů: role ekologických příležitostí a vyhynutí“. Paleobiologie. 42 (4): 547–573. doi:10.1017 / pab.2016.15. S2CID 3722056.
- ^ Kelley, N.P .; Motani, R. (2015). „Trofická konvergence pohání morfologickou konvergenci mořských tetrapodů“. Biologické dopisy. 11 (1): 20140709. doi:10.1098 / rsbl.2014.0709. PMC 4321144. PMID 25631228.
- ^ Fan, J.-X .; Shen, S.-Z .; Erwin, D.H .; Sadler, P.M .; MacLeod, N .; Cheng, Q.-M .; Hou, X.-D .; Yang, J .; Wang, X.-D .; Wang, Y .; Zhang, H .; Chen, X .; Li, G.-X .; Zhang, Y.-C .; Shi, Y.-K .; Yuan, D.-X .; Chen, Q .; Zhang, L.-N .; Li, C .; Zhao, Y.-Y. (2020). „Souhrn kambrijské až raně triasové biologické rozmanitosti mořských bezobratlých s vysokým rozlišením“. Věda. 367 (6475): 272–277. Bibcode:2020Sci ... 367..272F. doi:10.1126 / science.aax4953. PMID 31949075. S2CID 210698603.
- ^ Embry, A.F. (1997). „Globální hranice posloupnosti triasu a jejich identifikace v sedimentární pánvi západní Kanady“. Bulletin of Canadian Petroleum Geology. 45 (4): 415–433.
- ^ Paton, M. T.; Ivanov, A.V .; Fiorentini, M.L .; McNaughton, N.J .; Mudrovska, I .; Reznitskii, L.Z .; Demonterova, E.I. (2010). „Pozdní permské a raně triasové magmatické pulsy v synklině Angara – Taseeva, jižní sibiřské pasti a jejich možný vliv na životní prostředí“. Ruská geologie a geofyzika. 51 (9): 1012–1020. Bibcode:2010RuGG ... 51.1012P. doi:10.1016 / j.rgg.2010.08.009.
- ^ Takahashi, S .; Yamasaki, S.I .; Ogawa, K .; Kaiho, K .; Tsuchiya, N. (2015). „Redoxní podmínky na konci raného triasu Panthalassa“. Paleogeografie, paleoklimatologie, paleoekologie. 432: 15–28. Bibcode:2015PPP ... 432 ... 15T. doi:10.1016 / j.palaeo.2015.04.018.
- ^ Dick, D.G .; Maxwell, E.E. (2015). „Vývoj a zánik ichtyosaurů z pohledu kvantitativního modelování ekosystémů“. Biologické dopisy. 11 (7): 20150339. doi:10.1098 / rsbl.2015.0339. PMC 4528449. PMID 26156130.
- ^ Sander, P.M .; Chen, X .; Cheng, L .; Wang, X. (2011). „Krátkosrstý bezzubý ichtyosauř z Číny naznačuje pozdní triasovou diverzifikaci sání krmení ichthyosaurů“. PLOS ONE. 6 (5): e19480. Bibcode:2011PLoSO ... 619480S. doi:10.1371 / journal.pone.0019480. PMC 3100301. PMID 21625429.
- ^ Motani, R .; Ji, C .; Tomita, T .; Kelley, N .; Maxwell, E .; Jiang, D.-Y .; Sander, P.M. (2013). „Absence sání krmení ichthyosaurů a jeho důsledky pro triasovou mezopelagickou paleoekologii“. PLOS ONE. 8 (12): e66075. Bibcode:2013PLoSO ... 866075M. doi:10.1371 / journal.pone.0066075. PMC 3859474. PMID 24348983.
- ^ Vandewalle, P .; Saintin, P .; Chardon, M. (1995). "Struktury a pohyby bukálních a hltanových čelistí ve vztahu k podávání Diplodus sargus". Journal of Fish Biology. 46 (4): 623–656. doi:10.1111 / j.1095-8649.1995.tb01101.x.
- ^ Crofts, S.B .; Summers, A.P. (2014). „Jak nejlépe rozbít šneka: vliv tvaru zubu na drtivou zátěž“. Journal of the Royal Society Interface. 11 (92): 20131053. doi:10.1098 / rsif.2013.1053. PMC 3899874. PMID 24430124.
- ^ Mazouchova, N .; Umbanhowar, P.B .; Goldman, D.I. (2013). "Flipper-řízený pozemní pohyb robota inspirovaného mořskou želvou". Bioinspirace a biomimetika. 8 (2): 026007. Bibcode:2013BiBi ... 8b6007M. doi:10.1088/1748-3182/8/2/026007. PMID 23612858.
- ^ Fish, FE (2000). "Biomechanika a energetika u vodních a semiaquatických savců: ptakopysk až velryba". Fyziologická a biochemická zoologie. 73 (6): 683–698. doi:10.1086/318108. PMID 11121343.
- ^ Tytell, E.D .; Borazjani, I .; Sotiropoulos, F .; Baker, T.V .; Anderson, E.J .; Lauder, G.V. (2010). „Rozdělení funkčních rolí morfologie a pohybu při plavání ryb“. Integrativní a srovnávací biologie. 50 (6): 1140–1154. doi:10.1093 / icb / icq057. PMC 2981590. PMID 21082068.
- ^ Borazjani, I .; Sotiropoulos, F. (2010). "O roli formy a kinematiky na hydrodynamice plavání těla s vlastním pohonem / ocasní ploutví". Journal of Experimental Biology. 213 (1): 89–107. doi:10.1242 / jeb.030932. PMID 20008366. S2CID 27365252.
- ^ Gutarra, S .; Moon, B.C .; Rahman, I.A .; Palmer, C .; Lautenschlager, S .; Brimacombe, A.J .; Benton, M.J. (2019). „Účinky vývoje tělesného plánu na hydrodynamický odpor a energetické požadavky plavání v ichtyosaurech“. Sborník Královské společnosti B. 286 (1898): 20182786. doi:10.1098 / rspb.2018.2786. PMC 6458325. PMID 30836867.
- ^ A b Ji, C .; Tintori, A .; Jiang, D .; Motani, R. (2017). „New species of Thylacocephala (Arthropoda) from the Spathian (Lower Triassic) of Chaohu, Anhui Province of China“. PalZ. 91 (2): 171–184. doi:10.1007 / s12542-017-0347-7. S2CID 132045789.
- ^ A b Motani, R .; Jiang, D.-Y .; Tintori, A .; Ji, C .; Huang, J.-D. (2017). „Rozdíly mezozoických mořských plazů před a po masovém vyhynutí diktované časovou závislostí evolučních rychlostí“. Sborník Královské společnosti B. 284 (1854): 20170241. doi:10.1098 / rspb.2017.0241. PMC 5443947. PMID 28515201.
- ^ Fu, W .; Jiang, D .; Montañez, I .; Meylers, S .; Motani, R .; Tintori, A. (2016). „Cyklus uhlíku stimulovaný excentricitou a šikmostí v raném triasu a důsledky pro obnovu ekosystému po vyhynutí“. Vědecké zprávy. 6: 27793. Bibcode:2016NatSR ... 627793F. doi:10.1038 / srep27793. PMC 4904238. PMID 27292969.
- ^ Li, M .; Ogg, J .; Zhang, Y .; Huang, C .; Hinnov, L .; Chen, Z.Q .; Zou, Z. (2016). „Astronomické vyladění vyhynutí na konci permu a raná triasová epocha jižní Číny a Německa“. Dopisy o Zemi a planetách. 441: 10–25. Bibcode:2016E & PSL.441 ... 10L. doi:10.1016 / j.epsl.2016.02.017.
- ^ Ji, C .; Zhang, C .; Jiang, D.Y .; Bucher, H .; Motani, R .; Tintori, A. (2015). "Ammonoidní kontrola věku raných triasových mořských plazů z Chaohu (jižní Čína)". Palaeoworld. 24 (3): 277–282. doi:10.1016 / j.palwor.2014.11.009.
- ^ Li, S.Y .; Tong, J.N .; Liu, K.Y .; Wang, F.J .; Huo, Y.Y. (2007). "Lower Triassic cyklická depozice v Chaohu, provincie Anhui, Čína". Paleogeografie, paleoklimatologie, paleoekologie. 252 (1–2): 188–199. Bibcode:2007PPP ... 252..188S. doi:10.1016 / j.palaeo.2006.11.043.
- ^ Jiang, D.-Y .; Motani, R .; Tintori, A .; Rieppel, O .; Chen, G.-B .; Huang, J.-D .; Zhang, R .; Sun, Z.-Y .; Ji, C. (2014). „Počáteční triasový eosauropterygian Majiashanosaurus discocoracoidis, gen. et sp. listopad. (Reptilia, Sauropterygia), z Chaohu, provincie Anhui, Čínská lidová republika “. Časopis paleontologie obratlovců. 34 (5): 1044–1052. doi:10.1080/02724634.2014.846264. S2CID 129828376.
- ^ Sun, Z .; Tintori, A .; Jiang, D .; Motani, R. (2013). „New perleidid from the Spathian (Olenekian, Early Triassic) of Chaohu, Anhui Province, China“. Rivista Italiana di Paleontologia e Stratigrafia. 119 (3): 275–285. doi:10.13130/2039-4942/6040.
- ^ Tintori, A .; Huang, J.-D .; Jiang, D.-Y .; Sun, Z.-Y .; Motani, R .; Chen, G. (2014). „Nový Saurichthys (Actinopterygii) ze Spathian (raného triasu) z Chaohu (provincie Anhui, Čína) ". Rivista Italiana di Paleontologia e Stratigrafia. 120 (2): 157–164. doi:10.13130/2039-4942/6057.
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




