Agaróza - Agarose

Agaróza je polysacharid, obvykle extrahované z určitých červené mořské řasy.[1] Jedná se o lineární polymer složený z opakující se jednotky agarobiózy, což je a disacharid tvořeny D-galaktóza a 3,6-anhydro-L-galaktopyranóza.[2] Agaróza je jednou ze dvou hlavních složek agar, a je čištěn z agaru odstraněním další složky agaru, agaropektin.[3]
Agaróza se často používá v molekulární biologie pro separaci velkých molekul, zejména DNA tím, že elektroforéza. Desky z agarózových gelů (obvykle 0,7 - 2%) pro elektroforézu se snadno připraví nalitím teplého kapalného roztoku do formy. Široká škála různých agaróz různých molekulové hmotnosti a vlastnosti jsou pro tento účel komerčně dostupné. Agaróza může být také formována do perliček a použita v řadě chromatografické metody pro čištění bílkovin.
Struktura
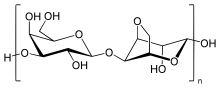
Agaróza je lineární polymer s molekulovou hmotností přibližně 120 000, který se skládá ze střídání D-galaktóza a 3,6-anhydro-L-galaktopyranóza spojená α- (1 → 3) a β- (1 → 4) glykosidickými vazbami. 3,6-anhydro-L-galaktopyranóza je L-galaktóza s anhydro můstkem mezi 3 a 6 pozicemi, i když některými L-galaktózové jednotky v polymeru nemusí obsahovat můstek. Nějaký D-galaktóza a L-galaktózové jednotky mohou být methylovaný, a pyruvát a síran se také nacházejí v malém množství.[4]
Každý agarózový řetězec obsahuje ~ 800 molekul galaktózy a polymerní řetězce agarózy tvoří spirálovitá vlákna, která se agregují do nadšroubovicové struktury s poloměrem 20-30nm.[5] Vlákna jsou kvazi-tuhá a mají široký rozsah délky v závislosti na koncentraci agarózy.[6] Po ztuhnutí tvoří vlákna a trojrozměrný síť kanálů o průměru v rozmezí od 50 nm do> 200 nm v závislosti na použité koncentraci agarózy - vyšší koncentrace poskytují nižší průměrné průměry pórů. 3D struktura je držena pohromadě s Vodíkové vazby a proto mohou být narušeny zahřátím zpět do kapalného stavu.
Vlastnosti
Agaróza je k dispozici ve formě bílého prášku, který se rozpouští v téměř vroucí vodě a po ochlazení vytvoří gel. Agaróza vykazuje fenomén termální hystereze ve svém přechodu kapalina na gel, tj. želatinuje a taje při různých teplotách. Teplota gelování a teplota tání se liší v závislosti na typu agarózy. Standardní agarózy odvozené z Gelidium má teplotu gelovatění 34–38 ° C (93–100 ° F) a teplotu tání 90–95 ° C (194–203 ° F), zatímco teplota tání Gracilaria, kvůli jeho vyšší methoxy substituenty, má teplotu gelovatění 40–52 ° C (104–126 ° F) a teplotu tání 85–90 ° C (185–194 ° F).[7] Teploty tání a gelovatění mohou záviset na koncentraci gelu, zejména při nízké koncentraci gelu nižší než 1%. Teploty želatinace a teploty tání jsou proto uvedeny při specifikované koncentraci agarózy.
Přírodní agaróza obsahuje nenabité methylové skupiny a rozsah methylace je přímo úměrný teplotě gelovatění. Syntetická methylace však má opačný účinek, přičemž zvýšená methylace snižuje teplotu gelovatění.[8] Chemickými modifikacemi je k dispozici celá řada chemicky modifikovaných agaróz s různými teplotami tání a gelovatění.
Agaróza v gelu tvoří síťovinu, která obsahuje póry, a velikost pórů závisí na koncentraci přidané agarózy. Při stání jsou agarózové gely náchylné k syneresis (vytlačování vody přes povrch gelu), ale proces je dostatečně pomalý, aby neinterferoval s použitím gelu.[9][10]
Agarózový gel může mít vysokou pevnost gelu při nízké koncentraci, takže je vhodný jako prostředek proti konvekci Gelová elektroforéza. Agarózové gely tak zředěné jako 0,15% mohou vytvářet desky pro gelovou elektroforézu.[11] Agarózový polymer obsahuje zejména nabité skupiny pyruvát a síran.[8] Tyto záporně nabité skupiny mohou zpomalit pohyb molekul DNA v procesu zvaném elektroendosmóza (EEO) a agaróza s nízkým EEO je proto obecně výhodná pro použití v agaróze gelová elektroforéza nukleových kyselin. K dispozici jsou také nulové EEO agarózy, které však mohou být pro některé aplikace nežádoucí, protože mohou být vyrobeny přidáním kladně nabitých skupin, které mohou ovlivnit následné enzymové reakce.[12] Elektroendosmóza je důvodem, proč se agaróza používá přednostně před agarem, protože agaropektin v agaru obsahuje významné množství negativně nabitých sulfátových a karboxylových skupin. Odstranění agaropektinu v agaróze podstatně snižuje EEO a také snižuje nespecifickou adsorpci biomolekul na gelovou matrici. Avšak pro některé aplikace, jako je elektroforéza sérových proteinů, může být žádoucí vysoký EEO a do použitého gelu může být přidán agaropektin.[13]
Agarózy s nízkou teplotou tání a gelovatění
Teploty tání a gelovatění agarózy mohou být modifikovány chemickými modifikacemi, nejčastěji hydroxyethylací, což snižuje počet vnitrodruhových vodíkových vazeb, což má za následek nižší teploty tání a nastavení než standardní agarózy.[14] Přesná teplota je určena stupněm substituce a mnoho dostupných agaróz s nízkým bodem tání (LMP) může zůstat tekutých v rozmezí 30–35 ° C (86–95 ° F). Tato vlastnost umožňuje enzymatický manipulace prováděné bezprostředně po elektroforéze na gelu DNA přidáním plátků roztaveného gelu obsahujícího požadovaný fragment DNA do reakční směsi. LMP agaróza obsahuje méně síranů, které mohou ovlivnit některé enzymatické reakce, a proto se s výhodou používá pro některé aplikace. Hydroxyethylace může snížit velikost pórů snížením hustoty balení agarózových svazků, proto může mít LMP gel také vliv na čas a separaci během elektroforézy.[15] Agarózy s extrémně nízkou teplotou tání nebo gelovatění mohou gelovat pouze při 8–15 ° C (46–59 ° F).
Aplikace

Agaróza je preferovaná matice pro práci s bílkoviny a nukleové kyseliny protože má širokou škálu fyzikální, chemické a tepelné stability a jeho nižší stupeň chemické složitosti také snižuje pravděpodobnost jeho interakce s biomolekuly. Agaróza se nejčastěji používá jako médium pro analytické měřítko elektroforetický oddělení v elektroforéza na agarózovém gelu. Gely vyrobené z purifikované agarózy mají relativně velkou velikost pórů, což je činí užitečnými pro separaci velkých molekul, jako jsou proteiny a proteinové komplexy> 200 kilodaltonů, stejně jako fragmenty DNA> 100 párů bází. Agaróza se také široce používá například pro řadu dalších aplikací imunodifúze a imunoelektroforéza, protože vlákna agarózy fungují jako kotva pro imunokomplexy.
Elektroforéza na agarózovém gelu
Rutinní metodou řešení je elektroforéza na agarózovém gelu DNA v laboratoři. Agarózové gely mají nižší rozlišovací schopnost pro DNA než akrylamidové gely, ale mají větší rozsah separace, a proto se obvykle používají pro fragmenty DNA o délce 50–20 000 bp (základní páry ), i když je možné rozlišení větší než 6 Mb gelová elektroforéza s pulzním polem (PFGE).[16] Může být také použit k oddělení velkých proteinových molekul a je to výhodná matrice pro gelovou elektroforézu částic s účinnou poloměry větší než 5-10 nm.[11]
Velikost pórů gelu ovlivňuje velikost DNA, kterou lze prosít. Čím nižší je koncentrace gelu, tím větší je velikost pórů a tím větší je DNA, kterou lze prosít. Gely s nízkou koncentrací (0,1 - 0,2%) jsou však křehké, a proto se s nimi obtížně manipuluje, a elektroforéza velkých molekul DNA může trvat několik dní. Mez rozlišení pro standardní elektroforézu na agarózovém gelu je kolem 750 kb.[16] Tuto hranici lze překonat pomocí PFGE, kde jsou na gel aplikována střídavá ortogonální elektrická pole. Fragmenty DNA se přeorientují, když aplikované pole změní směr, ale větším molekulám DNA trvá déle, než se znovu změní, když se změní elektrické pole, zatímco u menších je to rychlejší a DNA může být proto rozdělena podle velikosti.
Agarózové gely se odlévají do formy a po usazení obvykle běží vodorovně ponořené v pufrovacím roztoku. Tris-acetát-EDTA a Tris-borát-EDTA běžně se používají pufry, ale jiné pufry, jako je tris-fosfát, barbiturová kyselina, barbiturát sodný nebo tris-barbiturát vyrovnávací paměti lze použít v jiných aplikacích.[1] DNA je obvykle vizualizována barvením ethidiumbromid a poté zobrazeno pod a UV světlo, ale jsou k dispozici i jiné způsoby barvení, jako např SYBR Zelená, GelRed, methylenová modř, a křišťálově fialová. Pokud jsou oddělené fragmenty DNA potřebné pro další následný experiment, mohou být vyříznuty z gelu na plátky pro další manipulaci.

Čištění bílkovin
Agarózová gelová matrice se často používá pro čištění bílkovin například při separační přípravě na stupnici na koloně jako v gelová filtrační chromatografie, afinitní chromatografie a iontoměničová chromatografie. Nepoužívá se však jako kontinuální gel, spíše se formuje do porézních kuliček nebo pryskyřic různé jemnosti.[17] Korálky jsou vysoce porézní, takže bílkoviny mohou volně kuličkami protékat. Tyto perličky na bázi agarózy jsou obecně měkké a snadno se drtí, takže by se měly používat při gravitačním průtoku, nízkorychlostní centrifugaci nebo nízkotlakém postupu.[18] Pevnost pryskyřic lze zlepšit zvýšeným zesíťováním a chemickým vytvrzením agarózových pryskyřic, avšak tyto změny mohou také vést k nižší vazebné kapacitě proteinu v některých separačních postupech, jako je afinitní chromatografie.
Agaróza je užitečný materiál pro chromatografii, protože neabsorbuje biomolekuly ve významné míře, má dobré tokové vlastnosti a může tolerovat extrémy pH a iontová síla stejně jako vysoká koncentrace denaturanty například 8M močovina nebo 6M guanidin HCI.[19] Příklady matrice na bázi agarózy pro gelovou filtrační chromatografii jsou Sepharose a WorkBeads 40 SEC (zesítěná korálková agaróza), Praesto a Superose (vysoce zesítěné korálkové agarózy) a Superdex (dextran kovalentně navázaný na agarózu).
Pro afinitní chromatografii je perličková agaróza nejčastěji používanou matricovou pryskyřicí pro připojení ligandů, které vážou protein.[20] Ligandy jsou kovalentně spojeny přes spacer s aktivovanými hydroxylovými skupinami polymeru s agarózovými kuličkami. Zajímavé proteiny pak mohou být selektivně navázány na ligandy, aby se oddělily od ostatních proteinů, a poté je možné je eluovat. Použité agarózové kuličky mají typicky hustotu 4% a 6% s vysokou vazebnou kapacitou pro protein.
Pevná kultivační média
Agarózová destička může být někdy použita místo agaru pro kultivaci organismů, protože agar může obsahovat nečistoty, které mohou ovlivnit růst organismu, nebo některé následné postupy, jako je polymerázová řetězová reakce (PCR). Agaróza je také tvrdší než agar, a proto může být výhodnější, když je nutná větší pevnost gelu, a její nižší teplota želatinování může zabránit tepelný šok do organismu, když jsou buňky suspendovány v kapalině před gelovatěním. Může být použit pro kultivaci přísných autotrofních bakterií, rostlin protoplast,[21] Caenorhabditis elegans,[22] jiné organismy a různé buněčné linie.
3D buněčná kultura
Agaróza se často používá jako podpora trojrozměrné kultury lidí a zvířat buňky. Protože agaróza tvoří necytotoxický hydrogely, lze jej použít k reprodukci přirozeného prostředí buněk v lidském těle, extracelulární matrix. Agaróza však tvoří tuhou inertní látku hydrogel která nenese žádnou biologickou informaci, takže lidské a zvířecí buňky se k ní nemohou přilnout polysacharid. Kvůli těmto specifickým vlastnostem agaróza hydrogel napodobuje přirozené prostředí chrupavka buněk a bylo prokázáno, že podporují diferenciaci buněk chondrocyty do chrupavka. Za účelem úpravy mechanické vlastnosti agarózy k reprodukci přirozeného prostředí jiných lidských buněk může být agaróza chemicky modifikována přesnou oxidací primárního alkoholu D-galaktóza do karboxylová kyselina. Tato chemická modifikace poskytuje novou třídu materiálů zvanou karboxylovaná agaróza. Prostřednictvím kontroly nad počtem karboxylovaných D-galaktózy na polysacharid páteř, lze přesně řídit mechanické vlastnosti výsledného hydrogelu. Tyto karboxylované agarózové hydrogely se potom mohou kovalentně vázat peptidy za vzniku hydrogelu, na který mohou buňky přilnout. Ukázalo se, že tyto karboxylované agarózové hydrogely řídí organizaci lidských endotelových buněk do polarizovaných lumenů.[23]Smícháním plně karboxylované agarózy s přírodní agarózou lze připravit hydrogely, které pokrývají celou řadu mechanických vlastností.[24][25]
Testy motility
Někdy se místo agaru používá agaróza k měření motility a mobility mikroorganismů. Pohyblivé druhy budou schopny migrovat, i když pomalu, skrz porézní gel a poté lze vizualizovat míru infiltrace. Poréznost gelu přímo souvisí s koncentrací agaru nebo agarózy v médiu, takže k posouzení buněčné schopnosti lze použít různé koncentrační gely. plavání, rojení, klouzání a záškuby. Pro měření chemotaxe a chemokineze lze použít test migrace buněk pod agarózou. Mezi populaci buněk a a. Se umístí vrstva agarózového gelu chemoatraktant. Jak se koncentrační gradient vyvíjí z difúze chemoatraktantu do gelu lze následně vizualizovat různé buněčné populace vyžadující různé úrovně stimulace k migraci pomocí mikrofotografie, jak tunely nahoru skrz gel proti gravitaci podél gradientu.
Viz také
Reference
- ^ A b Jeppson, J. O .; C. B. Laurell; Bi Franzen (1979). „Elektroforéza na agarózovém gelu“. Klinická chemie. 25 (4): 629–638. doi:10.1093 / clinchem / 25.4.629. PMID 313856.
- ^ Agar Archivováno 16. října 2007, v Wayback Machine na lsbu.ac.uk Vodní struktura a věda
- ^ "Agar". Organizace pro výživu a zemědělství OSN.
- ^ Rafael Armisen; Fernando Galatas. „Kapitola 1 - Výroba, vlastnosti a použití agaru“. Fao.org.
- ^ Tom Maniatis; E. F. Fritsch; Joseph Sambrook (1982). "Kapitola 5, protokol 1". Molekulární klonování - laboratorní příručka. 1. p. 5.4. ISBN 978-0879691363.
- ^ Alistair M. Stephen; Glyn O. Phillips, eds. (2006). Potravinové polysacharidy a jejich aplikace. CRC Press. p. 226. ISBN 978-0824759223.
- ^ Workshop o biotechnologii mořských řas: souhrnná zpráva. Press National Academy. 1986. str. 25.
- ^ A b „Příloha B: Fyzikální chemie agarózy“ (PDF). Skupina Lonza.
- ^ S.E. Kopec; David A. Ledward; J. R. Mitchell, eds. (1998). Funkční vlastnosti potravinářských makromolekul. Springer. p. 149. ISBN 978-0-7514-0421-0.
- ^ Haesun Park; Kinam Park; Waleed S.W. Shalaby (1993). Biologicky odbouratelné hydrogely pro dodávání léčiv. CRC Press. p. 102. ISBN 978-1566760041.
- ^ A b Philip Serwer (1983). "Agarózové gely: Vlastnosti a použití pro elektroforézu". Elektroforéza. 4 (6): 375–382. doi:10,1002 / elps.1150040602. S2CID 97819634.
- ^ Joseph Sambrook; David Russell. "Kapitola 5, protokol 1". Molekulární klonování - laboratorní příručka. 1 (3. vyd.). p. 5.7. ISBN 978-0-87969-577-4.
- ^ Keren, David (26. září 2003). Elektroforéza bílkovin v klinické diagnostice. CRC Press. s. 7–8. ISBN 978-0340812136.
- ^ Tom Maniatis; E. F. Fritsch; Joseph Sambrook. "Kapitola 5, protokol 6". Molekulární klonování - laboratorní příručka. 1. p. 5.29. ISBN 978-0879695774.
- ^ Lee PY, Costumbrado J, Hsu CY, Kim YH (20. dubna 2012). „Elektroforéza na agarózovém gelu pro separaci fragmentů DNA“. J Vis Exp. 62 (62): 3923. doi:10.3791/3923. PMC 4846332. PMID 22546956.
- ^ A b Tom Maniatis; E. F. Fritsch; Joseph Sambrook (1982). "Kapitola 5, protokol 1". Molekulární klonování - laboratorní příručka. 1. p. 5.2–5.3. ISBN 978-0879691363.
- ^ David Freifelder (1982). Fyzikální biochemie: Aplikace na biochemii a molekulární biologii (2. vyd.). WH Freemane. p. 240. ISBN 978-0716714446.
- ^ "Přehled afinitního čištění". Thermo Scientific.
- ^ David Freifelder (1982). Fyzikální biochemie: Aplikace na biochemii a molekulární biologii (2. vyd.). WH Freemane. p. 258. ISBN 978-0716714446.
- ^ Pedro Cuatrecasas; Meir Wilchek (2004). William J. Lennarz; M. Daniel Lane (eds.). Encyclopedia of Biological Chemistry. Svazek 1. Akademický tisk. p. 52. ISBN 9780124437104.
- ^ J. M. Bonga; Patrick von Aderkas (1992). Kultura stromů in Vitro. Springer. p. 16. ISBN 978-0792315407.
- ^ Guy A. Caldwell; Shelli N. Williams; Kim A. Caldwell (2006). Integrovaná genomika: laboratorní kurz založený na objevech. Wiley. str. 94–95. ISBN 978-0470095027.
- ^ A. Zapomeňte; J. Christensen; S. Lüdeke; E. Kohler; S. Tobias; M. Matloubi; R. Thomann; V. P. Shastri (2013). „Polysacharidové hydrogely s nastavitelnou tuhostí a provaskulogenními vlastnostmi pomocí přepínače α-helix na β-list v sekundární struktuře“. Sborník Národní akademie věd Spojených států amerických. 110 (32): 12887–12892. Bibcode:2013PNAS..11212887F. doi:10.1073 / pnas.1222880110. PMC 3740890. PMID 23886665.
- ^ A. Zapomeňte; R. A. Pique; V. Ahamadi; S. Lüdeke; V. P. Shastri (2015). „Mechanicky přizpůsobené agarózové hydrogely prostřednictvím molekulárního legování s polysacharidy beta-listu“. Makromolekulární rychlá komunikace. 36 (2): 196–203. doi:10.1002 / marc.201400353. PMID 25250523.
- ^ A. Rüther; A. Zapomeňte; A. Roy; C. Carballo; F. Mießmer; R. K. Dukor; L. A. Nafie; C. Johannessen; V. P. Shastri; S. Lüdeke (2017). „odhalení přímé role polysacharidových beta-řetězců ve struktuře fyzikálních hydrogelů vyššího řádu“. Angewandte Chemie International Edition. 56 (16): 1–6. doi:10.1002 / anie.201701019. hdl:10067/1417890151162165141. PMID 28334501.