Rozpoznávání signálu částicové RNA - Signal recognition particle RNA - Wikipedia
| RN7SL1 | ||||||
|---|---|---|---|---|---|---|
| Identifikátory | ||||||
| Aliasy | RN7SL1, 7L1a, 7SL, RN7SL, RNSRP1, částice RNA pro rozpoznávání signálu, RNA, 7SL, cytoplazmatická 1, složka RNA částice pro rozpoznávání signálu 7SL1 | |||||
| Externí ID | OMIM: 612177 Genové karty: RN7SL1 | |||||
| Ortology | ||||||
| Druh | Člověk | Myš | ||||
| Entrez |
| |||||
| Ensembl |
| |||||
| UniProt |
|
| ||||
| RefSeq (mRNA) |
|
| ||||
| RefSeq (protein) |
|
| ||||
| Místo (UCSC) | n / a | n / a | ||||
| PubMed Vyhledávání | [1] | n / a | ||||
| Wikidata | ||||||

The signální rozpoznávací částicová RNA, (také známé jako 7SL, 6S, ffs nebo 4.5S RNA) je součástí částice pro rozpoznávání signálu (SRP) ribonukleoprotein komplex. SRP uznává signální peptid a váže se na ribozom, zastavení syntézy proteinů. SRP-receptor je protein zabudovaný do membrány, který obsahuje a transmembránový pór. Když SRP-ribozom komplex se váže na SRP-receptor„SRP uvolňuje ribozom a unáší se pryč. Ribozom obnovuje syntézu bílkovin, ale nyní se protein pohybuje přes SRP-receptor transmembránový pór.
Tímto způsobem SRP řídí pohyb bílkoviny v rámci buňka vázat se transmembránovými póry, které umožňují proteinu procházet membránou tam, kde je to potřeba. Složky RNA a bílkovin v tomto komplexu jsou vysoce konzervovaný ale liší se mezi různými království života.
Běžný SINUS rodina Alu pravděpodobně pochází z genu 7SL RNA po deleci centrální sekvence.[2]
The eukaryotický SRP se skládá z 300 nukleotidů 7S RNA a šest proteinů: SRP 72, 68, 54, 19, 14 a 9. Archaeal SRP se skládá z 7S RNA a homologů eukaryotických proteinů SRP19 a SRP54. Eukaryotické a archaální 7S RNA mají velmi podobné sekundární struktury.[3]
Ve většině bakterie, SRP sestává z molekuly RNA (4.5S) a proteinu Ffh (homolog eukaryotického proteinu SRP54). Nějaký Grampozitivní bakterie (např. Bacillus subtilis ) mají delší SRP RNA podobnou eukaryotům, která zahrnuje Alu doména.[4]
U eukaryot a archea se osm šroubovicových prvků vloží do Alu a S domény oddělené dlouhou linkerovou oblastí.[5][6] Má se za to, že Alu doména zprostředkovává retardační funkci SRP při prodloužení peptidového řetězce.[5] Univerzálně konzervovaná šroubovice, která interaguje s doménou SRP54 M, zprostředkovává rozpoznání signální sekvence.[6][7] Předpokládá se, že komplex SRP19-spirála 6 je zapojen do sestavování SRP a stabilizuje spirálu 8 pro SRP54. vazba[5] Lidé mají tři funkční geny SRP RNA, běžně pojmenované RN7SL1, RN7SL2 a RN7SL3. Zejména je známo, že lidský genom obsahuje velké množství sekvence související se SRP RNA, včetně Alu se opakuje.[4]
Objev
SRP RNA byla poprvé detekována u ptáků a myší onkogenní RNA (ocorna) virus částice.[8] Následně bylo zjištěno, že SRP RNA je stabilní složkou neinfikovaných buněk HeLa, kde je spojena s membrána a polysome zlomky.[9][10] V roce 1980 buněční biologové očistili od psí slinivka břišní 11S „protein rozpoznávající signál“ (náhodně také zkráceně „SRP“), který podporoval translokaci sekrečních proteinů přes membrána z endoplazmatické retikulum.[11] Poté bylo zjištěno, že SRP obsahoval RNA součástka.[12] Porovnání SRP RNA geny z různých druh odhalil, že šroubovice 8 SRP RNA je vysoce konzervovaný ve všech doménách život.[13] Oblasti v blízkosti 5'- a 3'konců savčí SRP RNA jsou podobné dominantní Alu rodině středních opakujících se sekvencí lidský genom.[14] Nyní se rozumí, že Alu DNA pochází ze SRP RNA excizí centrálního fragmentu specifického pro SRP RNA (S), po kterém následuje reverzní transkripce a integrace do více míst člověka chromozomy.[2] SRP RNA byly identifikovány také u některých organely, například v plastid SRP mnoha fotosyntetických organismů.[15]
Přepis a zpracování
Eukaryotické SRP RNA se přepisují DNA podle RNA polymeráza III (Pol III).[16] RNA polymeráza III také přepisuje geny na 5S ribozomální RNA, tRNA, 7SK RNA, a Spliceozomální RNA U6. The promotéři genů lidské SRP RNA zahrnuje prvky umístěné za transkripčním počátečním místem. Rostlina SRP RNA promotéři obsahovat upstream stimulační prvek (USE) a a TATA box.[Citace je zapotřebí ] Droždí Geny SRP RNA mají a TATA box a další intragenní promotorové sekvence (označované jako A- a B-bloky), které hrají roli při regulaci transkripce genu SRP Pol Pol III.[17] V bakterie, geny jsou organizovány v operony a přepsáno RNA polymeráza.[Citace je zapotřebí ] 5'-konec malé (4,5S) SRP RNA mnoha bakterií je štěpen RNáza P.[Citace je zapotřebí ] Konce Bacillus subtilis SRP RNA zpracovává RNáza III. Zatím žádná SRP RNA introny byly pozorovány.[Citace je zapotřebí ]
Funkce

Společná translokace
SRP RNA je nedílnou součástí malé a velké domény SRP. Funkce malé domény spočívá v oddálení translace proteinu, dokud nemá SRP vázaný na ribozomy příležitost spojit se s rezidentem membrány SRP receptor (SR). Ve velké doméně podporuje SRP RNA SRP nabitého signálním peptidem hydrolýzu dvou guanosin trifosfát (GTP) molekuly. Tato reakce uvolňuje SRP z receptoru SRP a ribozom, umožňující překlad pokračovat a protein vstoupit do translocon.[18] Protein transverzuje membránu kotranslačně (během translace) a vstupuje do jiného buněčného kompartmentu nebo extracelulárního prostoru. v eukaryoty, cílem je membrána z endoplazmatické retikulum (ER). v Archaea SRP dodává proteiny do plazmatická membrána.[19] V bakterie SRP primárně inkorporuje proteiny do vnitřní membrány.[20]
Posttranslační doprava
SRP se účastní také třídění bílkoviny po dokončení jejich syntézy (posttranslační třídění proteinů). v eukaryoty proteiny ukotvené v ocasu mající a hydrofobní inzertní sekvence na jejich C-konci jsou doručeny do endoplazmatické retikulum (ER) SRP.[21] Podobně SRP pomáhá posttranslačně při importu jaderně kódovaných proteinů do thylakoid membrána z chloroplasty.[22]
Struktura

V roce 2005, a nomenklatura pro všechny SRP RNA navrhovaný systém číslování 12 šroubovic. Sekce šroubovice jsou pojmenovány malým písmenem přípona (např. 5a). Vložkám nebo šroubovicím „větvím“ jsou dána tečkovaná čísla (např. 9.1 a 12.1).
SRP RNA je široká fylogenetické spektrum s ohledem na velikost a počet jeho strukturních znaků (viz příklady sekundární struktury SRP RNA níže). Nejmenší funkční SRP RNA byly nalezeny v mykoplazma a příbuzné druhy. Escherichia coli SRP RNA (také nazývaná 4.5S RNA) se skládá ze 114 nukleotid zbytky a tvoří RNA kmenová smyčka. The grampozitivní bakterie Bacillus subtilis kóduje větší 6S SRP RNA, která se podobá Archaeal homology ale chybí mu šroubovice SRP RNA 6. Archaeal SRP RNA mají šroubovice 1 až 8, chybí jim šroubovice 7 a jsou charakterizovány a terciární struktura který zahrnuje apikální smyčky šroubovice 3 a šroubovice 4. The eukaryotický SRP RNA postrádají spirálu 1 a obsahují spirálu 7 proměnné velikosti. Nějaký prvoky SRP RNA mají redukované šroubovice 3 a 4. The ascomycota SRP RNA mají zcela omezenou malou doménu a postrádají šroubovice 3 a 4. Největší dosud známé SRP RNA se nacházejí v droždí (Saccharomycetes ), který získal šroubovice 9 až 12 jako inzerce do šroubovice 5, stejně jako prodlouženou šroubovice 7. Semeno rostliny exprimují řadu vysoce divergentních SRP RNA.[3]
Motivy
Byly identifikovány čtyři konzervované znaky (motivy) (na obrázku jsou zobrazeny tmavě šedě): (1) vazebný motiv SRP54, (2) motiv Helralo 6 GNAR tetraloop, (3) motiv 5e a (4) UGU (NR) motiv.[Citace je zapotřebí ]
Vazba SRP54
Asymetrická smyčka mezi spirálovitými sekcemi 8a a 8b a sousední bází spárovanou sekcí 8b jsou prominentní vlastností každé SRP RNA. Čelní část 8b obsahuje ne-Watson-Crickovo párování bází které přispívají k tvorbě zploštělé malá drážka v RNA vhodné pro vazbu protein SRP54 (v bakteriích nazývaný Ffh).[6] The apikální smyčka šroubovice 8 obsahuje čtyři, pět nebo šest zbytků, v závislosti na druh. Má velmi konzervované guanosin jako první a adenosin jako poslední zbytek smyčky. Tato funkce je vyžadována pro interakci s třetí adenosin zbytek motivu tetraloop šroubovice 6 GNAR.[23]
Helix 6 GNAR tetraloop
SRP RNA z eukaryoty a Archaea mít GNAR tetraloop (N je pro všechny nukleotid, R je pro a purin ) ve šroubovici 6. Je konzervovaný adenosin zbytek je důležitý pro vazbu protein SRP19.[24] Tento adenosin provádí terciární interakci s jiným adenosin zbytky nacházející se v apikální smyčka šroubovice 8.[25]
5e
11 nukleotidů motivu 5e tvoří čtyři základní páry které jsou přerušeny smyčkou tří nukleotidy.[4] V eukaryoty, první nukleotid ve smyčce je adenosin který je potřebný pro vazbu protein SRP72.[26]
UGU (NR)
Motiv UGU (NR) spojuje šroubovice 3 a 4 v malé (Alu) doméně SRP. Houba SRP RNA bez helixů 3 a 4 obsahují motiv ve smyčce šroubovice 2.[4] Je to důležité ve vazbě SRP9 / 14 protein heterodimer jako součást RNA Otočit se.[27]
Sekundární
- Příklady sekundárních struktur SRP RNA
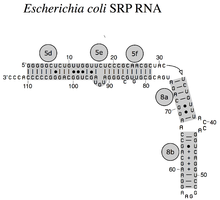
Bakteriální SRP RNA (4,5S RNA) z E-coli

Bakteriální SRP RNA (6S RNA) z Bacillus subtilis

Archaeal SRP RNA Archaeoglobus fulgidus

Eukaryotický protist SRP RNA z Trypanosoma brucei

Eukaryotický droždí SRP RNA z Saccharomyces cerevisiae





Terciární
| SRP RNA | |
|---|---|
| Identifikátory | |
| Rfam | CL00003 |
| Další údaje | |
| PDB struktur | PDBe 2IY3, 1Z43, 1RY1, 1QZW, 1MFQ, 1L9A, 1LNG,1JID, 1E8S, 1E8O, 1DUL, 1DUH,1D4R, 28SR, 28SP |
Rentgenová krystalografie, nukleární magnetická rezonance (NMR) a kryo-elektronová mikroskopie (cryo-EM] byly použity k určení molekulární struktura částí SRP RNA z různých druh. Dostupné struktury PDB ukazují, že molekula RNA je volná nebo pokud je navázána na jeden nebo více SRP bílkoviny.
- Krystalografické struktury reprezentativních SRP

SRP19-7S.S SRP RNA komplex z M. jannaschii[25]

S-doména lidského SRP[28]


Vazebné proteiny
Jeden nebo více SRP bílkoviny vázat na SRP RNA sestavit funkční SRP. SRP proteiny jsou pojmenovány podle jejich přibližné hodnoty molekulová hmotnost měřeno v kilodalton.[29] Většina bakteriální SRP se skládají ze SRP RNA a SRP54 (také nazývané Ffh pro „Fifty-Fnáš homolog ") Archaeal SRP obsahuje proteiny SRP54 a SRP19. v eukaryoty se SRP RNA kombinuje s importovanými proteiny SRP SRP9 / 14, SRP19 a SRP68 / 72 v oblasti jádro. Tento pre-SRP je transportován do cytosol kde se váže na protein SRP54.[30] The molekulární struktury volné nebo vázané na SRP RNA bílkoviny SRP9 / 14, SRP19 nebo SRP54 jsou známy ve vysokém rozlišení.
SRP9 a SRP14
SRP9 a SRP14 jsou strukturálně související a tvoří SRP9 / 14 heterodimer který se váže na SRP RNA malé (Alu) domény.[27] Droždí SRP postrádá SRP9 a obsahuje strukturálně příbuzný protein SRP21. Droždí SRP14 tvoří a homodimer.[31] SRP9 / 14 chybí v SRP z trypanosoma které místo toho vlastní a tRNA -jako molekula.[32]
SRP19
SRP19 se nachází v SRP z eukaryoty a Archaea. Jeho primární rolí je příprava SRP RNA pro vazbu SRP54, SRP68 a SRP72 správným uspořádáním šroubovic SRP RNA 6 a 8.[28] Droždí SRP obsahuje Sec65p, větší homolog SRP19.[33]
SRP54
Protein SRP54 (pojmenovaný Ffh v bakterie ) je základní součástí každého SRP. Skládá se ze tří funkčních domén: N-koncová (N) doména, GTPáza (G) doména a methionin -rich (M) doména.[34][35]
SRP68 a SRP72
Proteiny SRP68 a SRP72 jsou složkami velké domény eukaryotický SRP. Tvoří stabilní SRP68 / 72 heterodimer. Ukázalo se, že asi jedna třetina lidského proteinu SRP68 se váže na SRP RNA.[36] Relativně malá oblast umístěná blízko C-konce SRP72 se váže na motiv 5e SRP RNA.[26][37]
Reference
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ A b Ullu E, Tschudi C (1984). "Alu sekvence jsou zpracovány 7SL RNA geny". Příroda. 312 (5990): 171–172. Bibcode:1984Natur.312..171U. doi:10.1038 / 312171a0. PMID 6209580. S2CID 4328237.
- ^ A b Rosenblad MA, Larsen N, Samuelsson T, Zwieb C (2009). „Příbuznost v rodině SRP RNA“. RNA Biology. 6 (5): 508–516. doi:10,4161 / rna.6.5.9753. PMID 19838050.
- ^ A b C d Regalia M, Rosenblad MA, Samuelsson T (srpen 2002). "Predikce genů RNA pro rozpoznávání signálu". Výzkum nukleových kyselin. 30 (15): 3368–3377. doi:10.1093 / nar / gkf468. PMC 137091. PMID 12140321.
- ^ A b C Wild K, Weichenrieder O, Strub K, Sinning I, Cusack S (únor 2002). "Směrem ke struktuře částice pro rozpoznávání signálu savců". Aktuální názor na strukturní biologii. 12 (1): 72–81. doi:10.1016 / S0959-440X (02) 00292-0. PMID 11839493.
- ^ A b C Batey RT, Rambo RP, Lucast L, Rha B, Doudna JA (únor 2000). "Krystalová struktura ribonukleoproteinového jádra částice pro rozpoznávání signálu". Věda. 287 (5456): 1232–1239. Bibcode:2000Sci ... 287.1232B. doi:10.1126 / science.287.5456.1232. PMID 10678824.
- ^ Batey RT, Sagar MB, Doudna JA (březen 2001). "Strukturální a energetická analýza rozpoznávání RNA univerzálně konzervovaným proteinem z částice rozpoznávající signál". Journal of Molecular Biology. 307 (1): 229–246. doi:10.1006 / jmbi.2000.4454. PMID 11243816.
- ^ Bishop JM, Levinson WE, Sullivan D, Fanshier L, Quintrell N, Jackson J (prosinec 1970). „Nízkomolekulární RNA viru Rousova sarkomu. II. 7 S RNA“. Virologie. 42 (4): 927–937. doi:10.1016/0042-6822(70)90341-7. PMID 4321311.
- ^ Walker TA, Pace NR, Erikson RL, Erikson E, Behr F (září 1974). „7S RNA společná pro oncornaviry a normální buňky je spojena s polyribozomy“. Sborník Národní akademie věd Spojených států amerických. 71 (9): 3390–3394. Bibcode:1974PNAS ... 71,3390W. doi:10.1073 / pnas.71.9.3390. PMC 433778. PMID 4530311.
- ^ Zieve G, Penman S (květen 1976). "Malé druhy RNA buňky HeLa: metabolismus a subcelulární lokalizace". Buňka. 8 (1): 19–31. doi:10.1016/0092-8674(76)90181-1. PMID 954090. S2CID 26928799.
- ^ Walter P, Ibrahimi I, Blobel G (listopad 1981). „Translokace proteinů přes endoplazmatické retikulum. I. Protein pro rozpoznávání signálu (SRP) se váže na polysomy syntetizované sekreční protein in-vitro“. The Journal of Cell Biology. 91 (2 Pt 1): 545–550. doi:10.1083 / jcb.91.2.545. PMC 2111968. PMID 7309795.
- ^ Walter P, Blobel G (říjen 1982). „Částice pro rozpoznávání signálu obsahuje 7S RNA nezbytnou pro translokaci proteinu přes endoplazmatické retikulum“. Příroda. 299 (5885): 691–698. Bibcode:1982Natur.299..691W. doi:10.1038 / 299691a0. PMID 6181418. S2CID 4237513.
- ^ Larsen N, Zwieb C (leden 1991). "Zarovnání sekvence SRP-RNA a sekundární struktura". Výzkum nukleových kyselin. 19 (2): 209–215. doi:10.1093 / nar / 19.2.209. PMC 333582. PMID 1707519.
- ^ Ullu E, Murphy S, Melli M (květen 1982). „Lidská 7SL RNA se skládá ze 140 nukleotidové středně opakující se sekvence vložené do alu sekvence.“ Buňka. 29 (1): 195–202. doi:10.1016/0092-8674(82)90103-9. PMID 6179628. S2CID 12709599.
- ^ Rosenblad MA, Samuelsson T (listopad 2004). „Identifikace genů RNA pro rozpoznávání signálu chloroplastů“. Fyziologie rostlin a buněk. 45 (11): 1633–1639. doi:10.1093 / pcp / pch185. PMID 15574839.
- ^ Dieci G, Fiorino G, Castelnuovo M, Teichmann M, Pagano A (prosinec 2007). "Expandující transkriptom RNA RNA polymerázy III". Trendy v genetice. 23 (12): 614–622. doi:10.1016 / j.tig.2007.09.001. hdl:11381/1706964. PMID 17977614.
- ^ Dieci G, Giuliodori S, Catellani M, Percudani R, Ottonello S (březen 2002). „Intragenní adaptace promotoru a usnadnění recyklace RNA polymerázy III při transkripci SCR1, genu 7SL RNA Saccharomyces cerevisiae“. The Journal of Biological Chemistry. 277 (9): 6903–6914. doi:10,1074 / jbc.M105036200. PMID 11741971.
- ^ Shan SO, Walter P (únor 2005). "Cílení na ko-translační protein pomocí částice rozpoznávající signál". FEBS Dopisy. 579 (4): 921–926. doi:10.1016 / j.febslet.2004.11.049. PMID 15680975. S2CID 46046514.
- ^ Zwieb C, Eichler J (březen 2002). „Dostanete se na cíl: částice pro rozpoznávání signálu archaealu“. Archaea. 1 (1): 27–34. doi:10.1155/2002/729649. PMC 2685543. PMID 15803656.
- ^ Ulbrandt ND, Newitt JA, Bernstein HD (leden 1997). „Částice pro rozpoznávání signálu E. coli je nutná pro vložení podmnožiny proteinů vnitřní membrány“. Buňka. 88 (2): 187–196. doi:10.1016 / S0092-8674 (00) 81839-5. PMID 9008159. S2CID 15246619.
- ^ Abell BM, Pool MR, Schlenker O, Sinning I, High S (červenec 2004). „Částice pro rozpoznávání signálu zprostředkovává post-translační cílení u eukaryotů“. Časopis EMBO. 23 (14): 2755–2764. doi:10.1038 / sj.emboj.7600281. PMC 514945. PMID 15229647.
- ^ Schuenemann D, Gupta S, Persello-Cartieaux F, Klimyuk VI, Jones JD, Nussaume L, Hoffman NE (srpen 1998). „Nová částice pro rozpoznávání signálu se zaměřuje na proteiny sběru světla do tylakoidních membrán“. Sborník Národní akademie věd Spojených států amerických. 95 (17): 10312–10316. Bibcode:1998PNAS ... 9510312S. doi:10.1073 / pnas.95.17.10312. PMC 21505. PMID 9707644.
- ^ Zwieb C, van Nues RW, Rosenblad MA, Brown JD, Samuelsson T (leden 2005). „Nomenklatura pro všechny RNA částice rozpoznávající signál“. RNA. 11 (1): 7–13. doi:10,1261 / rna.7203605. PMC 1370686. PMID 15611297.
- ^ Zwieb C (srpen 1992). "Rozpoznání tetranukleotidové smyčky částice RNA rozpoznávající signál proteinem SRP19". The Journal of Biological Chemistry. 267 (22): 15650–15656. PMID 1379233.
- ^ A b Hainzl T, Huang S, Sauer-Eriksson AE (červen 2002). "Struktura komplexu SRP19 RNA a důsledky pro sestavu částice pro rozpoznávání signálu". Příroda. 417 (6890): 767–771. Bibcode:2002 Natur.417..767H. doi:10.1038 / nature00768. PMID 12050674. S2CID 2509475.
- ^ A b Iakhiaeva E, Wower J, Wower IK, Zwieb C (červen 2008). „Motiv 5e eukaryotické částice RNA pro rozpoznávání signálu obsahuje konzervovaný adenosin pro vazbu SRP72“. RNA. 14 (6): 1143–1153. doi:10,1261 / rna.979508. PMC 2390789. PMID 18441046.
- ^ A b Weichenrieder O, Wild K, Strub K, Cusack S (listopad 2000). "Struktura a sestavení Alu domény savčí částice pro rozpoznávání signálu". Příroda. 408 (6809): 167–173. Bibcode:2000Natur.408..167W. doi:10.1038/35041507. PMID 11089964. S2CID 4427070.
- ^ A b Kuglstatter A, Oubridge C, Nagai K (2002). "Indukované strukturální změny 7SL RNA během montáže částice pro rozpoznávání lidského signálu". Nat Struct Biol. 9 (10): 740–744. doi:10.1038 / nsb843. PMID 12244299. S2CID 9543041.
- ^ Walter P, Blobel G (září 1983). Msgstr "Demontáž a rekonstituce částice pro rozpoznávání signálu". Buňka. 34 (2): 525–533. doi:10.1016/0092-8674(83)90385-9. PMID 6413076. S2CID 17907778.
- ^ Politz JC, Yarovoi S, Kilroy SM, Gowda K, Zwieb C, Pederson T (leden 2000). "Součásti rozpoznávání signálu v jádře". Sborník Národní akademie věd Spojených států amerických. 97 (1): 55–60. Bibcode:2000PNAS ... 97 ... 55P. doi:10.1073 / pnas.97.1.55. PMC 26615. PMID 10618370.
- ^ Brooks MA, Ravelli RB, McCarthy AA, Strub K, Cusack S (květen 2009). "Struktura SRP14 z částice pro rozpoznávání signálu Schizosaccharomyces pombe". Acta Crystallographica oddíl D. 65 (Pt 5): 421–433. doi:10.1107 / S0907444909005484. PMID 19390147.
- ^ Liu L, Ben-Shlomo H, Xu YX, Stern MZ, Goncharov I, Zhang Y, Michaeli S (květen 2003). „Částice pro rozpoznávání signálu trypanosomatid se skládá ze dvou molekul RNA, homologu 7SL RNA a nové molekuly podobné tRNA“. The Journal of Biological Chemistry. 278 (20): 18271–18280. doi:10,1074 / jbc.M209215200. PMID 12606550.
- ^ Hann BC, Stirling CJ, Walter P (duben 1992). „Produkt genu SEC65 je podjednotkou částice rozpoznávající signál kvasinek, která je nutná pro její integritu.“ Příroda. 356 (6369): 532–533. Bibcode:1992 Natur.356..532H. doi:10.1038 / 356532a0. PMID 1313947. S2CID 4287636.
- ^ Römisch K, Webb J, Herz J, Prehn S, Frank R, Vingron M, Dobberstein B (srpen 1989). „Homologie 54K proteinu částice rozpoznávající signál, dokovacího proteinu a dvou proteinů E. coli s domnělými doménami vázajícími GTP“ (PDF). Příroda. 340 (6233): 478–482. Bibcode:1989 Natur.340..478R. doi:10.1038 / 340478a0. PMID 2502717. S2CID 4343347.
- ^ Bernstein HD, Poritz MA, Strub K, Hoben PJ, Brenner S, Walter P (srpen 1989). "Model pro rozpoznávání signální sekvence z aminokyselinové sekvence 54K podjednotky částice pro rozpoznávání signálu". Příroda. 340 (6233): 482–486. Bibcode:1989 Natur.340..482B. doi:10.1038 / 340482a0. PMID 2502718. S2CID 619959.
- ^ Iakhiaeva E, Bhuiyan SH, Yin J, Zwieb C (červen 2006). „Protein SRP68 částice rozpoznávající lidský signál: identifikace vazebných domén RNA a SRP72“. Věda o bílkovinách. 15 (6): 1290–1302. doi:10.1110 / ps.051861406. PMC 2242529. PMID 16672232.
- ^ Iakhiaeva E, Yin J, Zwieb C (leden 2005). "Identifikace domény vázající RNA v lidském SRP72". Journal of Molecular Biology. 345 (4): 659–666. doi:10.1016 / j.jmb.2004.10.087. PMID 15588816.
Další čtení
- Kuglstatter A, Oubridge C, Nagai K (říjen 2002). "Vyvolané strukturální změny 7SL RNA během montáže částice pro rozpoznávání lidského signálu". Přírodní strukturní biologie. 9 (10): 740–744. doi:10.1038 / nsb843. PMID 12244299. S2CID 9543041.
- Wang T, Tian C, Zhang W, Luo K, Sarkis PT, Yu L, Liu B, Yu Y, Yu XF (prosinec 2007). „7SL RNA zprostředkovává balení virionů antivirové cytidindeaminázy APOBEC3G“. Journal of Virology. 81 (23): 13112–13124. doi:10.1128 / JVI.00892-07. PMC 2169093. PMID 17881443.
- Ullu E, Weiner AM (prosinec 1984). "Lidské geny a pseudogeny pro 7SL RNA složku částice pro rozpoznávání signálu". Časopis EMBO. 3 (13): 3303–3310. doi:10.1002 / j.1460-2075.1984.tb02294.x. PMC 557853. PMID 6084597.
- Englert M, Felis M, Junker V, Beier H (prosinec 2004). „Nové upstreamové a intragenní kontrolní prvky pro transkripci lidských 7SL RNA genů závislou na RNA polymeráze III“. Biochimie. 86 (12): 867–874. doi:10.1016 / j.biochi.2004.10.012. PMID 15667936.
- Oubridge C, Kuglstatter A, Jovine L, Nagai K (červen 2002). "Krystalová struktura SRP19 v komplexu s S doménou SRP RNA a její důsledky pro sestavení částice pro rozpoznávání signálu". Molekulární buňka. 9 (6): 1251–1261. doi:10.1016 / S1097-2765 (02) 00530-0. PMID 12086622.
- Wang T, Tian C, Zhang W, Sarkis PT, Yu XF (leden 2008). „Interakce s 7SL RNA, ale nikoli s HIV-1 genomovou RNA nebo těly P, je nutná pro balení virionů APOBEC3F“. Journal of Molecular Biology. 375 (4): 1098–1112. doi:10.1016 / j.jmb.2007.11.017. PMID 18067920.
externí odkazy
- Databáze SRP (SRPDB): Zarovnání SRP RNA a souvisejících proteinů, sekundární struktury SRP RNA a 3-D modely
- Vstup Rfam pro částicovou RNA pro rozpoznávání signálu typu metazoanů
- Vstup Rfam pro bakteriální RNA s malým rozpoznáváním signálu
- Vstup Rfam pro bakteriální RNA s velkým rozpoznáváním signálu
- Vstup Rfam pro částicovou RNA pro rozpoznávání fungálního signálu
- Vstup Rfam pro částicovou RNA pro rozpoznávání signálu rostlin
- Vstup Rfam pro částicovou RNA pro rozpoznávání signálu protozoanem
- Vstup Rfam pro částicovou RNA pro rozpoznávání signálu Archaeal
- Částicový film pro rozpoznávání signálu Dnatube