Cortiho varhany - Organ of Corti
tento článek potřebuje další citace pro ověření. (Prosince 2014) (Zjistěte, jak a kdy odstranit tuto zprávu šablony) |
| Cortiho varhany | |
|---|---|
 Průřez hlemýžďem ilustrujícím Cortiho orgán | |
| Detaily | |
| Část | Kochlea z vnitřní ucho |
| Identifikátory | |
| latinský | organum spirale |
| Pletivo | D009925 |
| NeuroLex ID | birnlex_2526 |
| TA98 | A15.3.03.121 |
| TA2 | 7035 |
| FMA | 75715 |
| Anatomická terminologie | |
The varhany Cortinebo spirální orgán, je receptorový orgán pro sluch a nachází se v savci kochlea. Tento velmi pestrý pás epitelové buňky umožňuje přenos zvukových signálů do nervových impulsů akční potenciál.[1] K transdukci dochází prostřednictvím vibrací struktur ve vnitřním uchu, které způsobují posun kochleární tekutiny a pohyb vláskové buňky v Cortiho orgánu produkovat elektrochemické signály.[2]
Italský anatom Alfonso Giacomo Gaspare Corti (1822–1876) objevil Cortiho orgán v roce 1851.[3] Struktura se vyvinula z bazilární papila a je zásadní pro mechanotransdukce u savců.
Struktura

Cortiho orgán se nachází v Scala média z kochlea z vnitřní ucho mezi vestibulární potrubí a bubínku a je složen z mechanosenzorických buněk, známých jako vláskové buňky.[2] Strategicky umístěn na bazilární membrána Cortiho orgánu jsou tři řady vnější vlasové buňky (OHC) a jedna řada vnitřní vlasové buňky (IHC).[4] Oddělení těchto vlasových buněk podporuje buňky: Deitersovy buňky, také zvaný buňky falangy, které oddělují a podporují OHC i IHC.[4]
Vyčnívající z vrcholů vlasových buněk jsou malé prstové projekce zvané stereocilia, které jsou uspořádány odstupňovaným způsobem s nejkratší stereocilií na vnějších řadách a nejdelší ve středu. Tato gradace je považována za nejdůležitější anatomický rys Cortiho orgánu, protože to umožňuje smyslovým buňkám vynikající možnosti ladění.[5]
Pokud by byla hlemýžď rozvinutá, měla by délku asi 33 mm u žen a 34 mm u mužů, se standardní odchylkou pro populaci asi 2,28 mm.[6] Hlemýžď je také tonotopicky organizovaný, což znamená, že různé frekvence zvukových vln interagují s různými místy na struktuře. Základna hlemýžďů, nejblíže k vnějšímu uchu, je nejtuhší a nejužší a je místem, kde jsou přenášeny vysokofrekvenční zvuky. Vrchol nebo vrchol kochley je širší a mnohem pružnější a volnější a funguje jako místo transdukce nízkofrekvenčních zvuků.[7]
Funkce

Funkcí Cortiho orgánu je změnit (transduce ) sluchové signály a minimalizují extrakci zvukové energie vlasovými buňkami.[2] To je ušní boltec a střední ucho které fungují jako mechanické transformátory a zesilovače, takže zvukové vlny končí s amplitudami 22krát většími, než když vstoupily do ucha.
Sluchová transdukce
Za normálního sluchu pochází většina sluchových signálů, které se dostanou na první místo do Cortiho orgánu, z vnějšího ucha. Zvukové vlny vstoupit přes zvukovod a vibrovat ušní bubínek, také známý jako bubínek, který vibruje třemi malými kostmi zvanými ossicles. Ve výsledku je přiložen oválné okno se pohybuje a způsobí pohyb kulaté okno, což vede k posunutí kochleární tekutiny.[8]Ke stimulaci však může dojít také přímými vibracemi kochley z lebky. Ten druhý je označován jako Bone Conduction (nebo BC) sluch, který je doplňkem prvního popsaného, který se místo toho nazývá Air Conduction (nebo AC). AC i BC stimulují bazilární membránu stejným způsobem (Békésy, G.v., Experimental in Hearing. 1960).
Bazilární membrána na bubínku tlačí na vláskové buňky orgánu jako perilymfatický tlakové vlny procházejí. Stereocilie na vrcholu IHC se pohybuje s tímto posunem tekutin a v reakci na ně kation, nebo selektivní kladné ionty, kanály jsou taženy otevřenou kadherin struktury zvané odkazy na tipy které spojují sousední stereocilii.[9] Cortiho orgán obklopený tekutinou bohatou na draslík endolymfa, leží na bazilární membrána na základně Scala média. Pod Cortiho orgánem je scala tympani a nad ním scala vestibuli. Obě struktury existují v tekutině s nízkým obsahem draslíku perilymph.[8] Protože tyto stereocilie jsou uprostřed vysoké koncentrace draslíku, jakmile se jejich kationtové kanály otevřou, ionty draslíku i ionty vápníku proudí do horní části vlasové buňky. S tímto přílivem pozitivních iontů se stává IHC depolarizovaný, otevření napěťově řízených vápníkových kanálů v bazolaterální oblasti vlasových buněk a spuštění uvolnění neurotransmiteru glutamát. Elektrický signál je poté odeslán přes Sluchový nerv a do sluchová kůra mozku jako neurální zpráva.
Kochleární zesílení
Orgán Corti je také schopen modulovat sluchový signál.[7] Vnější vlasové buňky (OHC) mohou zesílit signál prostřednictvím tzv. Procesu elektromotilita kde zvyšují pohyb bazilárních a tectoriálních membrán a tím zvyšují vychýlení stereocilií v IHC.[8][10][11]
K tomu zásadní kus kochleární zesílení je motorický protein prestin, který mění tvar na základě napěťového potenciálu uvnitř vlasové buňky. Když je buňka depolarizována, prestin se zkrátí, a protože je umístěna na membráně OHC, natahuje bazilární membránu a zvyšuje její vychýlení, čímž vytváří intenzivnější účinek na vnitřní vlasové buňky (IHC). Když buňka hyperpolarizuje prestin, prodlužuje se a uvolňuje napětí na IHC, což snižuje nervové impulsy do mozku. Tímto způsobem je vlasová buňka sama schopná modifikovat sluchový signál, než se vůbec dostane do mozku.
Rozvoj
Orgán Corti, mezi scala tympani a Scala média, se vyvíjí po vzniku a růstu kochleární potrubí.[7] Vnitřní a vnější vlasové buňky se poté diferencují do příslušných pozic a jsou následovány organizací podpůrných buněk. Topologie podpůrných buněk se hodí pro skutečné mechanické vlastnosti, které jsou potřebné pro vysoce specializované pohyby vyvolané zvukem v Cortiho orgánu.[7]
Vývoj a růst Cortiho orgánu závisí na specifických genech, z nichž mnohé byly identifikovány v předchozím výzkumu (SOX2, GATA3, EYA1, FOXG1, BMP4, RAC1, a více),[7] podstoupit takovou diferenciaci. Konkrétně jde o růst kochleárního kanálu a tvorbu vláskových buněk v Cortiho orgánu.
Mutace v genech exprimovaných v Cortiho orgánu nebo v jeho blízkosti před diferenciací vláskových buněk budou mít za následek narušení diferenciace a potenciální nesprávnou funkci Cortiho orgánu.
Klinický význam
Ztráta sluchu
Cortiho orgán může být poškozen nadměrnou hladinou zvuku, což vede k poškození vyvolané hlukem.[12]
Nejběžnější druh sluchového postižení, senzorineurální ztráta sluchu, zahrnuje jako jednu z hlavních příčin snížení funkce v Cortiho orgánu. Konkrétně aktivní funkce zesílení vnější vlasové buňky je velmi citlivý na poškození způsobené traumatem z příliš hlasitých zvuků nebo na určité zvuky ototoxický léky. Jakmile jsou vnější vlasové buňky poškozené, neregenerují se a výsledkem je ztráta citlivosti a abnormálně vysoký nárůst hlasitosti (známý jako nábor) v části spektra, které slouží poškozeným buňkám.[13]
Zatímco ztráta sluchu byla u savců vždy považována za nevratnou, ryby a ptáci běžně takové poškození napravují. Studie z roku 2013 ukázala, že užívání konkrétních léků může reaktivovat geny, které se normálně exprimují pouze během vývoje vlasových buněk. Výzkum byl prováděn v Harvardská lékařská škola, Massachusetts oko a ucho a Keio University Lékařská fakulta v Japonsku.[14]
Další obrázky
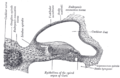
Příčný řez kochleárního kanálu fetální kočky.
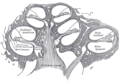
Schematický podélný řez hlemýžďem

Dno ductus cochlearis

Limbus laminæ spiralis a membrana basilaris
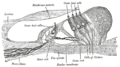
Řez spirálovým orgánem Corti (zvětšený)




Poznámky
- ^ Hudspeth, A (2014). "Integrace aktivního procesu vlasových buněk s kochleární funkcí". Recenze přírody Neurovědy. 15 (9): 600–614. doi:10.1038 / nrn3786. PMID 25096182. S2CID 3716179.
- ^ A b C Ucho Pujol, R., Irving, S., 2013
- ^ Betlejewski, S (2008). „Věda a život - historie markýze Alfonsa Cortiho“. Otolaryngologia Polska. 62 (3): 344–347. doi:10.1016 / S0030-6657 (08) 70268-3. PMID 18652163.
- ^ A b Malgrange, B; Van de Water, T. R.; Nguyen, L; Moonen, G; Lefebvre, P.P (2002). „Epiteliální podpůrné buňky se mohou diferencovat na vnější vlasové buňky a Deitersovy buňky v kultivovaném Cortiho orgánu“. Buněčné a molekulární biologické vědy. 59 (10): 1744–1757. doi:10.1007 / pl00012502. PMID 12475185. S2CID 2962483.
- ^ Lim, D (1986). "Funkční struktura Cortiho orgánu: recenze". Výzkum sluchu. 22 (1–3): 117–146. doi:10.1016/0378-5955(86)90089-4. PMID 3525482. S2CID 4764624.
- ^ Miller, J. D. (2007). „Pohlavní rozdíly v délce Cortiho orgánu u lidí“. The Journal of the Acoustical Society of America. 121 (4): EL151-5. Bibcode:2007ASAJ..121L.151M. doi:10.1121/1.2710746. PMID 17471760.
- ^ A b C d E Fritzsch, B; Jahan, já; Pan, N; Kers, J; Duncan, J; Kopecký, B (2012). „Rozptýlení molekulárního základu orgánu Cortiho vývoje: kde jsme teď?“. Výzkum sluchu. 276 (1–2): 16–26. doi:10.1016 / j.heares.2011.01.007. PMC 3097286. PMID 21256948.
- ^ A b C Nichols, J.G; Martin, A.R .; Fuchs, P.A .; Brown, D.A .; Diamond, ME; Weisblat, D.A (2012). Od Neurona po Brain, 5. vydání. Sunderland, MA: Sinauer Associates, Inc., str. 456–459. ISBN 978-0-87893-609-0.
- ^ Müller, Ulrich; Gillespie, Peter G .; Williams, David S .; Reynolds, Anna; Dumont, Rachel A .; Lillo, Concepcion; Siemens, leden (duben 2004). „Kadherin 23 je součástí koncovky ve stereocilii vlasových buněk“. Příroda. 428 (6986): 950–955. Bibcode:2004 Natur.428..950S. doi:10.1038 / nature02483. ISSN 1476-4687. PMID 15057245. S2CID 3506274.
- ^ Ashmore, Jonathan Felix (1987). „Rychlá pohybová odezva ve vnějších vlasových buňkách morčete: buněčný základ kochleárního zesilovače“. The Journal of Physiology. 388 (1): 323–347. doi:10.1113 / jphysiol.1987.sp016617. ISSN 1469-7793. PMC 1192551. PMID 3656195.

- ^ Ashmore, Jonathan (2008). „Kochleární pohyblivost vnějších vlasových buněk“. Fyziologické recenze. 88 (1): 173–210. doi:10.1152 / physrev.00044.2006. ISSN 0031-9333. PMID 18195086. S2CID 17722638.
- ^ Lim, David J. (březen 1986). "Účinky hluku a ototoxických léků na buněčné úrovni v hlemýždi: přehled". American Journal of Otolaryngology. 7 (2): 73–99. doi:10.1016 / S0196-0709 (86) 80037-0. PMID 3515985.
- ^ Robert A. Dobie (2001). Lékařsko-právní hodnocení ztráty sluchu. Thomson Delmar učení. ISBN 0-7693-0052-9.
- ^ „Cochlear hair cells - Beyond the Dish“. wordpress.com.
Reference
- Corti A (1851). „Recherches sur l'organe de l'ouïe des mammiferes“. Zeitschrift für Wissenschaftliche Zoologie. 3: 106–169.
- Fritzsch B, Jahan I, Pan N, Kersigo J, Duncan J, Kopecký B (2012). „Rozptýlení molekulárního základu orgánu Cortiho vývoje: kde jsme teď?“. Výzkum sluchu. 276 (1–2): 16–26. doi:10.1016 / j.heares.2011.01.007. PMC 3097286. PMID 21256948.
Dějiny. (n.d.).
- Hudspeth A (2014). "Integrace aktivního procesu vlasových buněk s kochleární funkcí". Recenze přírody Neurovědy. 15 (9): 600–614. doi:10.1038 / nrn3786. PMID 25096182. S2CID 3716179.
- Lim D (1986). "Funkční struktura Cortiho orgánu: recenze". Výzkum sluchu. 22 (1–3): 117–146. doi:10.1016/0378-5955(86)90089-4. PMID 3525482. S2CID 4764624.
- Malgrange B, Thiry M, Van de Water TR, Nguyen L, Moonen G, Lefebvre PP (2002). „Epiteliální podpůrné buňky se mohou diferencovat na vnější vlasové buňky a Deitersovy buňky v kultivovaném Cortiho orgánu“. Buněčné a molekulární biologické vědy. 59 (10): 1744–1757. doi:10.1007 / PL00012502. PMID 12475185. S2CID 2962483.
- Nicholls, J. G., Martin, A. R., Fuchs, P. A., Brown, D. A., Diamond, M. E., & Weisblat, D. A. (2012). Od Neurona k mozku (5. vydání, str. 456–459). Sunderland, MA: Sinauer Associates, Inc.
- Pritchard U. „Na Cortiho orgánu u savců“. 2. března 1876, Sborník královské společnosti v Londýně, svazek 24, str. 346–52 OCLC 1778190
- Pujol, R., & Irving, S. (2013). Ucho.
externí odkazy
- Disekující molekulární základ orgánu Cortiho vývoje PMC 3097286
- Varhany Cortiho 3D animace
- http://lobe.ibme.utoronto.ca/presentations/OHC_Electromotility/sld005.htm Schéma na University of Toronto
- http://mayoresearch.mayo.edu/mayo/research/ent_research/images/image02.gif Schéma na Mayo
- http://www.iurc.montp.inserm.fr/cric51/audition/english/corti/fcorti.htm na University of Montpellier 1
{kind=link}
| Kontrolní úřad |
|---|