Síť ve výchozím režimu - Default mode network
| Síť ve výchozím režimu | |
|---|---|
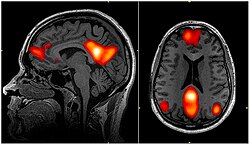 fMRI skenování zobrazení oblastí sítě ve výchozím režimu; the mediální prefrontální kůra, zadní cingulární kůra /precuneus a úhlový gyrus | |
| Anatomická terminologie |
v neurovědy, výchozí režim sítě (DMN), taky výchozí síťnebo výchozí stavová síť, je rozsáhlá mozková síť složený převážně z mediální prefrontální kůra, zadní cingulární kůra /precuneus a úhlový gyrus. To je nejlépe známé pro bytí aktivní, když člověk není zaměřen na vnější svět a mozek je na bdělý odpočinek, například během denní snění a bloudění mysli. Může být také aktivní během podrobných myšlenek souvisejících s výkonem externí úlohy.[3] Jindy, kdy je DMN aktivní, patří situace, kdy jedinec myslí na ostatní, myslí na sebe, vzpomíná na minulost a plánuje budoucnost.[4][5]
Ačkoli DMN byl původně si všiml, že je deaktivován u určitých úkolů zaměřených na cíl a je někdy označován jako negativní úkol síť,[6] může být aktivní v jiných cílově zaměřených úkolech, jako je sociální pracovní paměť nebo autobiografické úkoly.[7] Ukázalo se, že DMN negativně koreluje s jinými sítěmi v mozku, jako jsou sítě pozornosti.[8]
Důkazy poukazují na narušení DMN u lidí Alzheimerova choroba a poruchou autistického spektra.[4]
Dějiny
Hans Berger, vynálezce elektroencefalogram, jako první navrhl myšlenku, že mozek je neustále zaneprázdněn. V sérii prací publikovaných v roce 1929 ukázal, že elektrické oscilace detekované jeho zařízením nepřestávají, i když je subjekt v klidu. Jeho myšlenky však nebyly brány vážně a mezi neurology se vytvořilo obecné vnímání, že pouze když se provádí soustředěná činnost, aktivuje se mozek (nebo jeho část).[9]
Ale v padesátých letech minulého století si Louis Sokoloff a jeho kolegové všimli, že metabolismus v mozku zůstal stejný, když člověk přešel z klidového stavu do náročných matematických problémů, které naznačují, že aktivní metabolismus v mozku musí probíhat také během odpočinku.[4] V 70. letech David H. Ingvar a jeho kolegové pozorovali, že průtok krve v přední části mozku se stal nejvyšší, když je člověk v klidu.[4] Přibližně ve stejnou dobu, vnitřní oscilační chování v neuronech obratlovců bylo pozorováno v mozečku Purkyňovy buňky, nižší olivární jádro a thalamus.[10]
V 90. letech, s příchodem pozitronová emisní tomografie Skenování (PET) si vědci začali všímat, že když je člověk zapojen do úkolů vnímání, jazyka a pozornosti, stejné oblasti mozku se stávají méně aktivními než pasivní odpočinek, a označily tyto oblasti jako „deaktivované“.[4]
V roce 1995 Bharat Biswal, postgraduální student na lékařské fakultě ve Wisconsinu v Milwaukee, zjistil, že lidský senzomotorický systém vykazuje „konektivitu v klidovém stavu“ a vykazuje funkční synchronicitu magnetická rezonance (fMRI) skenuje, když není zapojen do žádného úkolu.[11][12]
Později experimenty od neurolog Marcus E. Raichle laboratoř v Lékařská fakulta Washingtonské univerzity a další skupiny [13] ukázaly, že mozková spotřeba energie se zvyšuje o méně než 5% jeho základní spotřeby energie při provádění soustředěného mentálního úkolu. Tyto experimenty ukázaly, že mozek je neustále aktivní s vysokou úrovní aktivity, i když se člověk nezabývá soustředěnou duševní prací. Výzkum se poté zaměřil na nalezení regionů odpovědných za tuto konstantní úroveň aktivity pozadí.[9]
Raichle vytvořil termín „výchozí režim“ v roce 2001 k popisu funkce mozku v klidovém stavu;[14] koncept se rychle stal ústředním tématem v roce 2006 neurovědy.[15] Kolem tentokrát byla vyvinuta myšlenka, že tato síť oblastí mozku je zapojena do vnitřně řízených myšlenek a je pozastavena během konkrétního chování zaměřeného na cíl. V roce 2003 Greicius a jeho kolegové zkoumali klidový stav fMRI skenuje a zkoumá, jak jsou vzájemně korelovány různé části mozku. Jejich korelační mapy zdůraznily stejné oblasti, které již identifikovali ostatní vědci.[16] To bylo důležité, protože to demonstrovalo sbližování metod, které všechny vedly k zapojení stejných oblastí do DMN. Od té doby další klidové státní sítě Byly nalezeny (RSN), jako jsou vizuální, sluchové a sítě pozornosti. Některé z nich často korelují se sítí ve výchozím režimu.[8]

Na začátku až v polovině 2000s vědci označili síť ve výchozím režimu jako záporná síť úkolu[6] protože to bylo deaktivováno, když účastníci museli plnit úkoly. DMN se považovalo za aktivní pouze během pasivního odpočinku a poté se vypnulo během externě zaměřených úkolů zaměřených na cíl. Studie však prokázaly, že DMN je aktivní v externích cílených úkolech, o nichž je známo, že zahrnují DMN, jako je sociální pracovní paměť nebo autobiografické úkoly.[7] DMN také ukazuje vyšší aktivaci, když jsou behaviorální reakce stabilní, a tato aktivace je nezávislá na bloudění mysli nahlášené samy sebou.[17]
Kolem roku 2007 vzrostl počet článků odkazujících na síť ve výchozím režimu.[18] Ve všech letech před rokem 2007 vyšlo 12 článků, které v názvu odkazovaly na „síť ve výchozím režimu“ nebo „výchozí síť“; mezi lety 2007 a 2014 se však počet zvýšil na 1 384 prací. Jedním z důvodů pro zvýšení počtu článků je výsledek robustního efektu nalezení DMN se skenováním v klidovém stavu a analýza nezávislých komponent (ICA).[13][19] Dalším důvodem je, že DMN lze měřit pomocí krátkých a snadných skenů v klidovém stavu, což znamená, že je lze provádět u jakékoli populace včetně malých dětí, klinických populací a primátů jiných než lidských.[4] Třetím důvodem je, že role DMN byla rozšířena na více než jen na pasivní mozkovou síť.[Citace je zapotřebí ]
Funkce
Síť ve výchozím režimu je považována za součást několika různých funkcí:
Je to potenciálně neurologický základ pro já:[18]
- Autobiografické informace: Vzpomínky na sbírku událostí a faktů o sobě samém
- Vlastní reference: Mluvit o vlastnostech a popisech vlastního já
- Emoce sebe sama: Úvahy o vlastním emocionálním stavu
Přemýšlím o ostatních:[18]
- Teorie mysli: Přemýšlet o myšlenkách druhých a o tom, co mohou nebo nemusí vědět
- Emoce jiných: Pochopení emocí druhých lidí a vcítění se do jejich pocitů
- Morální uvažování: Určení spravedlivého a nespravedlivého výsledku akce
- Sociální hodnocení: Úsudky o špatném přístupu k sociálním konceptům
- Sociální kategorie: Úvahy o důležitých sociálních charakteristikách a postavení skupiny
Vzpomínka na minulost a přemýšlení o budoucnosti:[18]
- Vzpomínka na minulost: Připomínáme si události, které se staly v minulosti
- Představujeme si budoucnost: Představujeme si události, které by se mohly v budoucnu stát
- Epizodická paměť: Podrobná paměť související s konkrétními událostmi v čase
- Pochopení příběhu: Pochopení a zapamatování si příběhu
Síť ve výchozím režimu je aktivní během pasivního odpočinku a bloudění mysli[4] což obvykle zahrnuje přemýšlet o druhých, přemýšlet o sobě, vzpomínat na minulost a představovat si budoucnost, spíše než vykonaný úkol.[18] Nedávná práce však zpochybnila konkrétní mapování mezi sítí ve výchozím režimu a putováním myslí, vzhledem k tomu, že systém je důležitý při udržování podrobných reprezentací informací o úkolu během kódování pracovní paměti.[20] Elektrokortikografie studie (zahrnující umisťování elektrod na povrch pokožky hlavy) ukázaly, že síť ve výchozím režimu se aktivuje za zlomek sekundy poté, co účastníci dokončí úkol.[21] Navíc se během úkolů náročných na pozornost ukázalo, že dostatečná deaktivace sítě ve výchozím režimu v době kódování paměti vede k úspěšnější dlouhodobé konsolidaci paměti.[22]
Studie ukázaly, že když lidé sledují film,[23] poslouchat příběh,[24][25] nebo si přečtěte příběh[26] jejich DMN jsou navzájem vysoce korelované. DMN nesouvisí, pokud jsou příběhy zakódovány nebo jsou v jazyce, kterému člověk nerozumí, což naznačuje, že síť je silně zapojena do porozumění a následné formování paměti tohoto příběhu.[25] Ukázalo se, že DMN dokonce souvisí, pokud je stejný příběh prezentován různým lidem v různých jazycích,[27] dále naznačuje, že DMN je skutečně zapojen do aspektu porozumění příběhu, nikoli do sluchového nebo jazykového aspektu.
Ukázalo se, že síť ve výchozím režimu se deaktivuje během externích úkolů zaměřených na cíl, jako je vizuální pozornost nebo kognitivní funkce pracovní paměť úkoly, což vede některé výzkumníky k tomu, aby označili síť jako záporná síť.[6] Pokud jsou však úkoly externími cílovými úkoly, o nichž je známo, že jsou rolí DMN, jako je sociální pracovní paměť nebo autobiografický úkol, je DMN s úkolem pozitivně aktivován a koreluje s jinými sítěmi, jako je síť v výkonná funkce.[7]
Dosud netušenou možností je, že výchozí síť je aktivována imobilizací, která je vlastní zkušební proceduře (pacient je připoután na zádech na nosítkách a vložen úzkým tunelem do masivní kovové konstrukce). Tento postup vytváří pocit zachycení a není překvapením, že nejčastěji uváděným vedlejším účinkem je klaustrofobie. Tento alternativní pohled navrhuje nedávný článek, který odkazuje teorie mysli k imobilizaci.[28]
Nedávný výzkum ukázal, že DMN souvisí s vnímáním krása, ve kterém se síť aktivuje zobecněným způsobem do esteticky se pohybujících domén, jako jsou umělecká díla, krajina a architektura. To by vysvětlovalo hluboký vnitřní pocit rozkoše související s estetika, propojené s pocitem osobní identity, kvůli síťovým funkcím souvisejícím s já.[29]
Anatomie
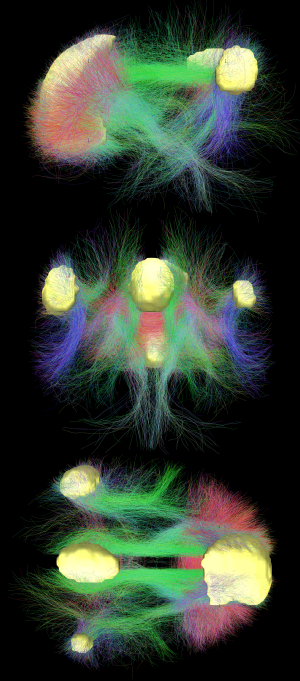
Síť výchozího režimu je vzájemně propojená a anatomicky definovaná[4] soubor oblastí mozku. Síť lze rozdělit na rozbočovače a podsekce:
Funkční náboje:[31] Informace o sobě
- Zadní cingulární kůra (PCC) & precuneus: Kombinuje pozornost zdola nahoru (neřízenou) s informacemi z paměti a vnímání. Ventrální (spodní) část PCC se aktivuje ve všech úkolech, které zahrnují DMN, včetně těch, které se týkají já, vztahují se k ostatním, pamatují si minulost, přemýšlejí o budoucnosti a zpracovávají koncepty plus prostorovou navigaci. Hřbetní (horní) část PCC zahrnuje nedobrovolné vědomí a vzrušení. Precuneus je zapojen do vizuálních, senzomotorických a pozorovacích informací.
- Mediální prefrontální kůra (mPFC): Rozhodnutí o vlastním zpracování, jako jsou osobní informace, autobiografické vzpomínky, budoucí cíle a události, a rozhodování o osobně velmi blízkých osobách, jako je rodina. Ventrální (dolní) část je zapojena do pozitivních emocionálních informací a interně oceňované odměny.
- Úhlový gyrus: Spojuje vnímání, pozornost, prostorové poznání a akci a pomáhá s částmi vzpomínek na epizodické vzpomínky
Hřbetní mediální subsystém:[31] Myslet na ostatní
- Funkční náboje: PCC, mPFC, a úhlový gyrus
- Hřbetní mediální prefrontální kůra (dmPFC): Podílí se na sociálně zaměřeném myšlení, jako je určování nebo odvozování účelu akcí druhých
- Dočasné spojení (TPJ): Odráží přesvědčení o ostatních, také známé jako teorie mysli
- Laterální temporální kůra: Získávání sociálních sémantický a koncepční znalosti
- Přední časový pól: Abstraktní koncepční informace zvláště sociální povahy
Mediální dočasný subsystém:[31] Autobiografické paměť a budoucí simulace
- Funkční náboje: PCC, mPFC, a úhlový gyrus
- Hippocampus (HF +): Formování nových vzpomínek, vzpomínání na minulost a představování budoucnosti
- Parahippocampus (PHC): Prostorové a scénické rozpoznávání a simulace
- Retrospleniální kůra (RSC): Prostorová navigace
- Zadní dolní mozkový lalok (pIPL): Spojení sluchových, vizuálních a somatosenzorických informací a pozornosti
Síť výchozího režimu je nejčastěji definována pomocí klidový stav data vložením semene do zadní cingulární kůra a zkoumání, které další oblasti mozku nejvíce korelují s touto oblastí.[16] DMN lze také definovat podle oblastí deaktivovaných během externích řízených úkolů ve srovnání s odpočinkem.[14] Analýza nezávislých komponent (ICA) robustně najde DMN pro jednotlivce i napříč skupinami a stal se standardním nástrojem pro mapování výchozí sítě.[13][19]
Ukázalo se, že síť ve výchozím režimu vykazuje nejvyšší přesah ve své strukturální a funkční konektivitě, což naznačuje, že strukturální architektura mozku může být postavena takovým způsobem, že je tato konkrétní síť ve výchozím nastavení aktivována.[1] Nedávné důkazy ze studie populačního zobrazování mozku 10 000 účastníků britské biobanky dále naznačují, že každý uzel DMN lze rozložit na podoblasti s doplňkovými strukturálními a funkčními vlastnostmi. Ve výzkumu DMN bylo rozšířenou praxí považovat jeho uzly, které tvoří, za funkčně homogenní, ale rozdíl mezi poduzly v každém hlavním uzlu DMN byl většinou zanedbáván. Blízká blízkost subnodů, které šíří hipokampální časoprostorové výstupy, a subnodů, které popisují architekturu globální sítě, však může umožnit výchozí funkce, jako je autobiografické vyvolání nebo interně orientované myšlení.[32]
V mozku kojenců existují omezené důkazy o výchozí síti, ale výchozí síťové připojení je u dětí ve věku 9–12 let konzistentnější, což naznačuje, že výchozí síť prochází vývojovými změnami.[8]
Analýza funkční konektivity u opic ukazuje podobnou síť regionů jako síť ve výchozím režimu, kterou vidíme u lidí.[4] PCC je také klíčovým centrem u opic; mPFC je však menší a méně dobře propojený s jinými oblastmi mozku, hlavně proto, že lidský mPFC je mnohem větší a dobře vyvinutý.[4]
Difúzní MRI zobrazovací show bílá hmota plochy spojující různé oblasti DMN dohromady.[18] Strukturální spojení zjištěná difuzním zobrazením MRI a funkční korelace z klidový stav fMRI vykazují nejvyšší úroveň překrývání a shody v oblastech DMN.[1] To poskytuje důkaz, že neurony v oblastech DMN jsou navzájem spojeny prostřednictvím velkých ploch axonů, což způsobuje vzájemnou korelaci aktivity v těchto oblastech.
Patofyziologie
Předpokládá se, že síť výchozího režimu je relevantní pro poruchy včetně Alzheimerova choroba, autismus, schizofrenie, Deprese, chronická bolest, posttraumatická stresová porucha a další.[4][33] Zejména DMN bylo také hlášeno, že vykazuje překrývající se, přesto odlišné vzorce nervové aktivity napříč různými podmínkami duševního zdraví, například při přímém porovnání porucha pozornosti s hyperaktivitou a autismus.[34]
Lidé s Alzheimerovou chorobou vykazují snížení hladiny glukózy (spotřeby energie) v oblastech sítě výchozího režimu.[4] Tato snížení začínají jako mírná snížení u mírných pacientů a pokračují ve velkých sníženích u těžkých pacientů. Překvapivě narušení DMN začínají ještě předtím, než jednotlivci projeví známky Alzheimerovy choroby.[4] Pozemky peptidu amyloid-beta, o nichž se předpokládá, že způsobují Alzheimerovu chorobu, ukazují, že tvorba peptidu je v DMN.[4] To přimělo Randyho Bucknera a kolegy navrhnout vysokou rychlost metabolismu z nepřetržité aktivace DMN, což způsobí, že se v těchto oblastech DMN hromadí více amyloid-beta peptidu.[4] Tyto amyloid-beta peptidy narušují DMN a protože DMN je silně zapojen do tvorby a získávání paměti, vede toto narušení k symptomům Alzheimerovy choroby.
DMN je považován za narušený u jedinců s poruchou autistického spektra.[4][35] Tito jedinci jsou narušeni v sociální interakci a komunikaci, což jsou úkoly ústřední v této síti. Studie prokázaly horší souvislosti mezi oblastmi DMN u jedinců s autismem, zejména mezi mPFC (zapojeným do myšlení o sobě a ostatních) a PCC (centrální jádro DMN).[36][37] Čím závažnější je autismus, tím méně jsou tyto oblasti navzájem propojeny.[36][37] Není jasné, zda je to příčina nebo důsledek autismu, nebo zda třetí faktor způsobuje obojí (matoucí ).
Nižší konektivita mezi oblastmi mozku byla zjištěna napříč výchozí sítí u lidí, kteří zažili dlouhodobé trauma, jako je zneužívání nebo zanedbávání v dětství, a je spojena s dysfunkčními příloha vzory. Mezi lidmi zažívajícími posttraumatická stresová porucha, byla nalezena nižší aktivace v zadním cingulárním gyrusu ve srovnání s kontrolami a závažná PTSD byla charakterizována nižší konektivitou v DMN.[33][38] Hyperkonektivita výchozí sítě byla spojena s přežvykováním při depresi první epizody[39] a chronická bolest.[40]Pokud se změní síť ve výchozím režimu, může to změnit způsob, jakým člověk vnímá události a jejich sociální a morální úvahy, čímž se člověk stane náchylnějším k velkým depresivním příznakům.[41]
Multivariační analýza odhalila genetické asociace klidové DMN u psychotické bipolární poruchy a schizofrenie.[42]
Modulace
Síť výchozího režimu (DMN) může být modulovaný následujícími zásahy a procesy:
- Akupunktura - Deaktivace limbických oblastí mozku a DMN.[43] Bylo navrženo, že je to kvůli reakci na bolest.[44]
- Rozjímání - Strukturální změny v oblastech DMN, jako je temporoparietální spojení, zadní cingulární kůra, a precuneus byly nalezeny u odborníků na meditaci.[45] U dlouhodobých lékařů dochází ke snížení aktivace a snížení funkční konektivity DMN.[45] Různé formy nedirektivní meditace, včetně Transcendentální meditace[46] a Acem meditace,[47] Bylo zjištěno, že aktivují DMN.
- Spící a odpočinkovou bdělost
- Klidová bdělost - Funkční propojení mezi uzly DMN je silné.[48]
- Nástup spánku - Snížení připojení mezi DMN a síť pozitivních úkolů.[48]
- Fáze N2 spánku NREM - Snížení konektivity mezi zadní cingulární kůra a mediální prefrontální kůra.[48]
- Fáze N3 spánku NREM - Další pokles připojení mezi PCC a MPFC.[48]
- REM spánek - Možné zvýšení konektivity mezi uzly DMN.[48]
- Nedostatek spánku - Funkční konektivita mezi uzly DMN v klidovém stavu je obvykle silná, ale deprivace spánku vede k poklesu konektivity v rámci DMN.[49] Nedávné studie naznačují snížení konektivity mezi DMN a sítí pozitivních úkolů v důsledku ztráty spánku.[50]
- Psychedelické léky - Snížený průtok krve do PCC a mPFC byl pozorován při podávání psilocybin. Tyto dvě oblasti jsou považovány za hlavní uzly DMN.[51] Jedna studie o účincích látky LSD prokázáno, že lék desynchronizuje mozkovou aktivitu v DMN; aktivita oblastí mozku, které tvoří DMN, bude méně korelována.[52]
- Hluboká stimulace mozku - Změny v mozkové činnosti s hlubokou mozkovou stimulací lze použít k vyvážení klidových stavových sítí.[53]
- Psychoterapie - V PTSD se abnormality v síti výchozího režimu normalizují u jedinců, kteří reagují na psychoterapeutické intervence.[54][55]
- Antidepresiva - Změny v připojení DMN se snižují po léčbě antidepresivy v PTSD.[55]
- Fyzická aktivita a cvičení - Fyzická aktivita a pravděpodobnější Aerobní trénink, může změnit DMN. Kromě toho sportovní odborníci ukazují rozdíly v sítích, zejména pokud jde o DMN.[56][57][58]
Kritika
Někteří argumentovali, že mozkové oblasti v síti se standardním režimem se zobrazují společně pouze kvůli vaskulární vazbě velkých tepen a žil v mozku v blízkosti těchto oblastí, ne proto, že jsou tyto oblasti ve skutečnosti navzájem funkčně spojeny. Podpora tohoto argumentu pochází ze studií, které ukazují, že změna dechu mění hladinu kyslíku v krvi, což zase nejvíce ovlivňuje DMN.[4] Tyto studie však nevysvětlují, proč lze DMN identifikovat také pomocí PET skenuje měřením metabolismu glukózy, který je nezávislý na vazbě cév[4] a v elektrokortikografie studie[59] měření elektrické aktivity na povrchu mozku a v MEG měřením magnetických polí spojených s elektrofyziologickou mozkovou aktivitou, která obchází hemodynamickou odpověď.[60]
Myšlenka „výchozí sítě“ není všeobecně přijímána.[61] V roce 2007 byl koncept výchozího režimu kritizován jako nepoužitelný pro pochopení funkce mozku, protože jednodušší hypotéza spočívá v tom, že klidový mozek ve skutečnosti provádí více zpracování než mozek provádějící určité „náročné“ úkoly a že neexistuje zvláštní význam pro vnitřní aktivitu klidového mozku.[62]
Viz také
- Funkční zobrazování magnetickou rezonancí (fMRI)
- Klidový stav fMRI
- Putování mysli
- Záporná síť úkolu
- Síť kladná na úkoly
Reference
- ^ A b C Horn, Andreas; Ostwald, Dirk; Reisert, Marco; Blankenburg, Felix (2013). „Strukturálně-funkční konektom a síť výchozího režimu lidského mozku“. NeuroImage. 102: 142–151. doi:10.1016 / j.neuroimage.2013.09.069. PMID 24099851.
- ^ Garrity, A .; Pearlson, G. D .; McKiernan, K .; Lloyd, D .; Kiehl, K. A .; Calhoun, V. D. (2007). "Aberantní výchozí režim funkční konektivity u schizofrenie". Dopoledne. J. Psychiatrie. 164 (3): 450–457. doi:10.1176 / ajp.2007.164.3.450. PMID 17329470.
- ^ Sormaz, Mladen; Murphy, Charlotte; Wang, Hao-Ting; Hymers, Mark; Karapanagiotidis, Theodoros; Poerio, Giulia; Margulies, Daniel S .; Jefferies, Elizabeth; Smallwood, Jonathan (2018). „Síť ve výchozím režimu může podporovat úroveň podrobností během aktivních stavů úkolů“. Sborník Národní akademie věd. 115 (37): 9318–9323. doi:10.1073 / pnas.1721259115. PMC 6140531. PMID 30150393.
- ^ A b C d E F G h i j k l m n Ó p q r Buckner, R.L .; Andrews-Hanna, J. R .; Schacter, D. L. (2008). „Výchozí síť mozku: anatomie, funkce a význam pro nemoc“. Annals of the New York Academy of Sciences. 1124 (1): 1–38. Bibcode:2008NYASA1124 ... 1B. CiteSeerX 10.1.1.689.6903. doi:10.1196 / annals.1440.011. PMID 18400922.
- ^ Lieberman, Matthew (2. září 2016). Sociální. Broadway Books. p. 19. ISBN 978-0-307-88910-2.
- ^ A b C Fox, Michael D .; Snyder, Abraham Z .; Vincent, Justin L .; Corbetta, Maurizio; Van Essen, David C .; Raichle, Marcus E. (05.07.2005). „Lidský mozek je vnitřně organizován do dynamických antikorelujících funkčních sítí.“. Sborník Národní akademie věd Spojených států amerických. 102 (27): 9673–9678. Bibcode:2005PNAS..102,9673F. doi:10.1073 / pnas.0504136102. ISSN 0027-8424. PMC 1157105. PMID 15976020.
- ^ A b C Spreng, R. Nathan (01.01.2012). „Klam„ záporné úlohy “sítě. Hranice v psychologii. 3: 145. doi:10.3389 / fpsyg.2012.00145. ISSN 1664-1078. PMC 3349953. PMID 22593750.
- ^ A b C Broyd, Samantha J .; Demanuele, Charmaine; Debener, Stefan; Pomáhá, Suzannah K .; James, Christopher J .; Sonuga-Barke, Edmund J. S. (2009). „Dysfunkce mozku ve výchozím režimu u duševních poruch: systematický přehled“. Neurovědy a biobehaviorální recenze. 33 (3): 279–96. doi:10.1016 / j.neubiorev.2008.09.002. PMID 18824195.
- ^ A b Raichle, Marcus (březen 2010). „Mozková temná energie“. Scientific American. 302 (3): 44–49. Bibcode:2010SciAm.302c..44R. doi:10.1038 / scientificamerican0310-44. PMID 20184182.
- ^ Llinas, R. R. (2014). „Vnitřní elektrické vlastnosti savčích neuronů a funkce CNS: historická perspektiva“. Neurosci přední buňky. 8: 320. doi:10.3389 / fncel.2014.00320. PMC 4219458. PMID 25408634.
- ^ Biswal, B; Yetkin, F. Z .; Haughton, V. M .; Hyde, J. S. (1995). "Funkční konektivita v motorické kůře klidového lidského mozku pomocí echoplanarové MRI". Magn Reson Med. 34 (4): 537–541. doi:10,1002 / mrm.1910340409. PMID 8524021. S2CID 775793.
- ^ Shen, H. H. (2015). „Základní koncepty: Připojení v klidovém stavu“. Sborník Národní akademie věd. 112 (46): 14115–14116. Bibcode:2015PNAS..11214115S. doi:10.1073 / pnas.1518785112. PMC 4655520. PMID 26578753.
- ^ A b C Kiviniemi, Vesa J .; Kantola, Juha-Heikki; Jauhiainen, Jukka; Hyvärinen, Aapo; Tervonen, Osmo (2003). "Analýza nezávislých komponent nedeterministických zdrojů signálu fMRI". NeuroImage. 19 (2 Pt 1): 253–260. doi:10.1016 / S1053-8119 (03) 00097-1. PMID 12814576.
- ^ A b Raichle, M. E.; MacLeod, A. M .; Snyder, A. Z .; Powers, W. J .; Gusnard, D. A .; Shulman, G.L. (2001). „Inaugurační článek: Výchozí režim funkce mozku“. Sborník Národní akademie věd. 98 (2): 676–82. Bibcode:2001PNAS ... 98..676R. doi:10.1073 / pnas.98.2.676. PMC 14647. PMID 11209064.
- ^ Raichle, Marcus E .; Snyder, Abraham Z. (2007). "Výchozí režim funkce mozku: Stručná historie vyvíjejícího se nápadu". NeuroImage. 37 (4): 1083–90. doi:10.1016 / j.neuroimage.2007.02.041. PMID 17719799.
- ^ A b Greicius, Michael D .; Krasnow, Ben; Reiss, Allan L .; Menon, Vinod (01.01.2003). „Funkční konektivita v klidovém mozku: síťová analýza hypotézy výchozího režimu“. Sborník Národní akademie věd Spojených států amerických. 100 (1): 253–258. Bibcode:2003PNAS..100..253G. doi:10.1073 / pnas.0135058100. ISSN 0027-8424. PMC 140943. PMID 12506194.
- ^ Kucyi, Aaron (2016). „Spontánní výchozí síťová aktivita odráží variabilitu chování nezávisle na putování myslí“. PNAS. 113 (48): 13899–13904. doi:10.1073 / pnas.1611743113. PMID 27856733.
- ^ A b C d E F Andrews-Hanna, Jessica R. (06.06.2012). „Výchozí síť mozku a jeho adaptivní role ve vnitřní mentaci“. The Neuroscientist: A Review Journal Bringing Neurobiology, Neurology and Psychiatry. 18 (3): 251–270. doi:10.1177/1073858411403316. ISSN 1089-4098. PMC 3553600. PMID 21677128.
- ^ A b De Luca, M; Beckmann, CF; De Stefano, N; Matthews, PM; Smith, SM (2006-02-15). „Sítě klidového stavu fMRI definují odlišné způsoby interakcí na dálku v lidském mozku“. NeuroImage. 29 (4): 1359–1367. doi:10.1016 / j.neuroimage.2005.08.035. PMID 16260155.
- ^ Sormaz, Mladen; Murphy, Charlotte; Wang, Ha-ting; Hymers, Mark; Karapanagiotidis, Theodoros; Poerio, Giulia; Margulies, Daniel S .; Jefferies, Elizabeth; Smallwood, Jonathan (2018-08-24). „Síť ve výchozím režimu může podporovat aktivní úroveň podrobností během aktivních stavů úkolů“. Sborník Národní akademie věd. 115 (37): 9318–9323. doi:10.1073 / pnas.1721259115. ISSN 0027-8424. PMC 6140531. PMID 30150393.
- ^ Dastjerdi, Mohammad; Foster, Brett L .; Nasrullah, Sharmin; Rauschecker, Andreas M .; Dougherty, Robert F .; Townsend, Jennifer D .; Chang, Catie; Greicius, Michael D .; Menon, Vinod (2011-02-15). „Diferenciální elektrofyziologická odezva během odpočinku, autoreferenční a nereferenční úkoly v lidské posteromediální kůře“. Sborník Národní akademie věd Spojených států amerických. 108 (7): 3023–3028. Bibcode:2011PNAS..108.3023D. doi:10.1073 / pnas.1017098108. ISSN 1091-6490. PMC 3041085. PMID 21282630.
- ^ Lefebvre, Etienne; D’Angiulli, Amedeo (2019). „Verbální učení zprostředkované snímky závisí na interakcích živosti a známosti: možná role interference činnosti sítě v dualistickém klidovém stavu“. Vědy o mozku. 9 (6): 143. doi:10,3390 / brainsci9060143. ISSN 2076-3425. PMC 6627679. PMID 31216699.
- ^ Hasson, Uri; Furman, Orit; Clark, Dav; Dudai, Yadin; Davachi, Lila (07.02.2008). „Vylepšené korelace mezi objekty během sledování filmu korelují s úspěšným epizodickým kódováním“. Neuron. 57 (3): 452–462. doi:10.1016 / j.neuron.2007.12.009. ISSN 0896-6273. PMC 2789242. PMID 18255037.
- ^ Lerner, Yulia; Zlato, Christopher J .; Silbert, Lauren J .; Hasson, Uri (2011-02-23). „Topografické mapování hierarchie časových receptivních oken pomocí vyprávěného příběhu“. The Journal of Neuroscience. 31 (8): 2906–2915. doi:10.1523 / JNEUROSCI.3684-10.2011. ISSN 1529-2401. PMC 3089381. PMID 21414912.
- ^ A b Simony, Erez; Zlato, Christopher J; Chen, Janice; Lositsky, Olga; Yeshurun, Yaara; Wiesel, Ami; Hasson, Uri (2016-07-18). "Dynamická rekonfigurace sítě ve výchozím režimu během narativního porozumění". Příroda komunikace. 7 (1): 12141. Bibcode:2016NatCo ... 712141S. doi:10.1038 / ncomms12141. ISSN 2041-1723. PMC 4960303. PMID 27424918.
- ^ Regev, Mor; Zlato, Christopher J .; Simony, Erez; Hasson, Uri (02.10.2013). „Selektivní a neměnné nervové reakce na mluvené a psané příběhy“. The Journal of Neuroscience. 33 (40): 15978–15988. doi:10.1523 / JNEUROSCI.1580-13.2013. ISSN 1529-2401. PMC 3787506. PMID 24089502.
- ^ Zlato, Christopher J .; Thompson, Christopher R .; Lerner, Yulia; Hasson, Uri (2012-10-31). „Neztrácí se v překladu: nervové odpovědi sdílené napříč jazyky“. The Journal of Neuroscience. 32 (44): 15277–15283. doi:10.1523 / JNEUROSCI.1800-12.2012. ISSN 1529-2401. PMC 3525075. PMID 23115166.
- ^ Tsoukalas, Ioannis (2017). „Theory of Mind: Towards the Evolutionary Theory“. Evoluční psychologická věda. 4: 38–66. doi:10.1007 / s40806-017-0112-x.Pdf.
- ^ Starr, G. Gabrielle; Stahl, Jonathan L .; Belfi, Amy M .; Isik, Ayse Ilkay; Vessel, Edward A. (2019-09-04). „Síť ve výchozím režimu představuje estetickou přitažlivost, která se zobecňuje napříč vizuálními doménami“. Sborník Národní akademie věd. 116 (38): 19155–19164. doi:10.1073 / pnas.1902650116. ISSN 0027-8424. PMC 6754616. PMID 31484756.
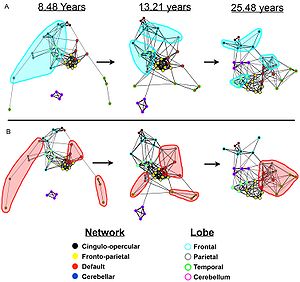
- ^ Fair, Damien A .; Cohen, Alexander L .; Moc, Jonathan D .; Dosenbach, Nico U. F .; Church, Jessica A .; Miezin, Francis M .; Schlaggar, Bradley L .; Petersen, Steven E. (2009). Sporns, Olaf (ed.). „Funkční mozkové sítě se vyvíjejí z„ místní distribuované “organizace“. PLOS výpočetní biologie. 5 (5): e1000381. Bibcode:2009PLSCB ... 5E0381F. doi:10.1371 / journal.pcbi.1000381. PMC 2671306. PMID 19412534.
- ^ A b C Andrews-Hanna, Jessica R .; Smallwood, Jonathan; Spreng, R. Nathan (01.05.2014). „Výchozí síť a myšlenka generovaná samy: procesy komponent, dynamická kontrola a klinická relevance“. Annals of the New York Academy of Sciences. 1316 (1): 29–52. Bibcode:2014NYASA1316 ... 29A. doi:10.1111 / nyas.12360. ISSN 1749-6632. PMC 4039623. PMID 24502540.
- ^ Kernbach, J.M .; Yeo, B.T.T .; Smallwood, J .; Margulies, D.S .; Thiebaut; de Schotten, M .; Walter, H .; Sabuncu, M.R .; Holmes, A.J .; Gramfort, A .; Varoquaux, G .; Thirion, B .; Bzdok, D. (2018). „Subspecializace v rámci standardních uzlů charakterizovaných 10 000 účastníky britské biobanky“. Proc. Natl. Acad. Sci. 115 (48): 12295–12300. doi:10.1073 / pnas.1804876115. PMC 6275484. PMID 30420501.
- ^ A b Akiki, Teddy J .; Averill, Christopher L .; Wrocklage, Kristen M .; Scott, J. Cobb; Averill, Lynnette A .; Schweinsburg, Brian; Alexander-Bloch, Aaron; Martini, Brenda; Southwick, Steven M .; Krystal, John H .; Abdallah, Chadi G. (2018). „Abnormality sítě ve výchozím režimu u posttraumatické stresové poruchy: nový přístup topologie omezený na síť“. NeuroImage. 176: 489–498. doi:10.1016 / j.neuroimage.2018.05.005. ISSN 1053-8119. PMC 5976548. PMID 29730491.
- ^ Kernbach, Julius M .; Satterthwaite, Theodore D .; Bassett, Danielle S .; Smallwood, Jonathan; Margulies, Daniel; Krall, Sarah; Shaw, Philip; Varoquaux, Gaël; Thirion, Bertrand; Konrad, Kerstin; Bzdok, Danilo (17. července 2018). „Sdílené endo-fenotypy dysfunkce výchozího režimu u poruchy pozornosti / hyperaktivity a poruchy autistického spektra“. Translační psychiatrie. 8 (1): 133. doi:10.1038 / s41398-018-0179-6. PMC 6050263. PMID 30018328.
- ^ Vigneshwaran S, Mahanand B. S., Suresh S, Sundararajan N. Identifikace rozdílů v mozkových činnostech a přesná detekce poruchy autistického spektra pomocí zobrazování funkční magnetické rezonance v klidovém stavu: Přístup prostorové filtrace. Lékařská analýza obrazu. 2017;35:375–389. doi:10.1016 / j.media.2016.08.003.
- ^ A b Washington, Stuart D .; Gordon, Evan M .; Brar, Jasmit; Warburton, Samantha; Sawyer, Alice T .; Wolfe, Amanda; Mease-Ference, Erin R .; Girton, Laura; Hailu, Ayichew (01.04.2014). "Dysmaturace sítě ve výchozím režimu u autismu". Mapování lidského mozku. 35 (4): 1284–1296. doi:10,1002 / hbm.22252. ISSN 1097-0193. PMC 3651798. PMID 23334984.
- ^ A b Yerys, Benjamin E .; Gordon, Evan M .; Abrams, Danielle N .; Satterthwaite, Theodore D .; Weinblatt, Rachel; Jankowski, Kathryn F .; Strang, John; Kenworthy, Lauren; Gaillard, William D. (01.01.2015). „Výchozí segregace v síti a sociální deficity u poruchy autistického spektra: důkazy od neléčivých dětí“. NeuroImage: Klinické. 9: 223–232. doi:10.1016 / j.nicl.2015.07.018. PMC 4573091. PMID 26484047.
- ^ Dr. Ruth Lanius, konference Brain Mapping, Londýn, listopad 2010
- ^ Zhu, X; Wang, X; Xiao, J; Liao, J; Zhong, M; Wang, W; Yao, S (2012). „Důkaz o disociačním vzoru v síťovém připojení v klidovém stavu ve výchozím režimu u pacientů s první depresí, kteří dosud nebyli léčeni, s depresí.“ Biologická psychiatrie. 71 (7): 611–7. doi:10.1016 / j.biopsych.2011.10.035. PMID 22177602.
- ^ Kucyi, A; Moayedi, M; Weissman-Fogel, I; Goldberg, M. B .; Freeman, B. V .; Tenenbaum, H. C .; Davis, K. D. (2014). „Vylepšená mediální prefrontální-výchozí síťová funkční konektivita v chronické bolesti a její asociace s přežvykováním bolesti“. Journal of Neuroscience. 34 (11): 3969–75. doi:10.1523 / JNEUROSCI.5055-13.2014. PMC 6705280. PMID 24623774.
- ^ Sambataro, Fabio; Vlk, Nadine; Giusti, Pietro; Vasic, Nenad; Vlk, Robert (říjen 2013). „Síť ve výchozím režimu v depresi: Cesta k narušenému afektivnímu poznávání?“ (PDF). Klinická neurální psychologie. 10: 212–216. Archivovány od originál (PDF) dne 29. srpna 2017. Citováno 28. září 2017.
- ^ Meda, Shashwath A .; Ruaño, Gualberto; Windemuth, Andreas; O’Neil, Kasey; Berwise, Clifton; Dunn, Sabra M .; Boccaccio, Leah E .; Narayanan, Balaji; Kocherla, Mohan (2014-05-13). „Vícerozměrná analýza odhaluje genetické asociace sítě klidového výchozího režimu u psychotické bipolární poruchy a schizofrenie“. Sborník Národní akademie věd. 111 (19): E2066 – E2075. Bibcode:2014PNAS..111E2066M. doi:10.1073 / pnas.1313093111. ISSN 0027-8424. PMC 4024891. PMID 24778245.
- ^ Huang, Wenjing; Pach, Daniel; Napadow, Vitaly; Park, Kyungmo; Long, Xiangyu; Neumann, Jane; Maeda, Yumi; Nierhaus, Till; Liang, Fanrong; Witt, Claudia M .; Harrison, Ben J. (9. dubna 2012). „Charakterizace akupunkturních stimulů pomocí zobrazování mozku pomocí fMRI - systematický přehled a metaanalýza literatury“. PLOS ONE. 7 (4): e32960. Bibcode:2012PLoSO ... 732960H. doi:10.1371 / journal.pone.0032960. PMC 3322129. PMID 22496739.
- ^ Chae, Younbyoung; Chang, Dong-Seon; Lee, Soon-Ho; Jung, Won-Mo; Lee, In-Seon; Jackson, Stephen; Kong, Jian; Lee, Hyangsook; Park, Hi-Joon; Lee, Hyejung; Wallraven, Christian (březen 2013). „Inserting Needles Into the Body: a Meta-Analysis of Brain Activity associated with Acupuncture Needle Stimulation“. The Journal of Pain. 14 (3): 215–222. doi:10.1016 / j.jpain.2012.11.011. PMID 23395475.
- ^ A b Fox, Kieran C. R .; Nijeboer, Savannah; Dixon, Matthew L .; Floman, James L .; Ellamil, Melissa; Rumak, Samuel P .; Sedlmeier, Peter; Christoff, Kalina (2014). „Je meditace spojena se změněnou strukturou mozku? Systematický přehled a metaanalýza morfometrického neuroobrazu u lékařů“. Neurovědy a biobehaviorální recenze. 43: 48–73. doi:10.1016 / j.neubiorev.2014.03.016. PMID 24705269.
- ^ Raffone, Antonino; Srinivasan, Narayanan (2010). „Zkoumání meditace v neurovědě pozornosti a vědomí“. Kognitivní zpracování. 11 (1): 1–7. doi:10.1007 / s10339-009-0354-z. PMID 20041276.
- ^ Xu, J; Vik, A; Groote, IR; Lagopoulos, J; Holen, A; Ellingsen, Ø; Håberg, AK; Davanger, S (2014). „Nedirektivní meditace aktivuje výchozí síť a oblasti spojené s načítáním paměti a emočním zpracováním.“. Přední. Hučení. Neurosci. 8 (86): 86. doi:10.3389 / fnhum.2014.00086. PMC 3935386. PMID 24616684.
- ^ A b C d E Picchioni, Dante; Duyn, Jeff H .; Horovitz, Silvina G. (2013-10-15). "Spánek a funkční konektom". NeuroImage. 80: 387–396. doi:10.1016 / j.neuroimage.2013.05.067. PMC 3733088. PMID 23707592.
- ^ McKenna, Benjamin S .; Eyler, Lisa T. (2012). „Překrývající se prefrontální systémy zapojené do kognitivního a emočního zpracování u euthymické bipolární poruchy a následující deprivace spánku: Přehled funkčních neuroimagingových studií“. Recenze klinické psychologie. 32 (7): 650–663. doi:10.1016 / j.cpr.2012.07.003. PMC 3922056. PMID 22926687.
- ^ Basner, Mathias; Rao, Hengyi; Goel, Namni; Dinges, David F (říjen 2013). „Spánková deprivace a neurobehaviorální dynamika“. Aktuální názor v neurobiologii. 23 (5): 854–863. doi:10.1016 / j.conb.2013.02.008. PMC 3700596. PMID 23523374.
- ^ Carhart-Harris, Robin L .; Erritzoe, David; Williams, Tim; Stone, James M .; Reed, Laurence J .; Colasanti, Alessandro; Tyacke, Robin J .; Leech, Robert; Malizia, Andrea L .; Murphy, Kevin; Hobden, Peter; Evans, John; Feilding, Amanda; Wise, Richard G .; Nutt, David J. (2012). „Neurální koreláty psychedelického stavu, jak je stanoveno ve studiích fMRI s psilocybinem“. PNAS. 109 (6): 2138–2143. doi:10.1073 / pnas.1119598109. PMC 3277566. PMID 22308440.
- ^ Carhart-Harris, Robin L .; Muthukumaraswamy, Suresh; Roseman, Leor; Kaelen, Mendel; Droog, Wouter; Murphy, Kevin; Tagliazucchi, Enzo; Schenberg, Eduardo E .; Nest, Timothy; Orban, Csaba; Leech, Robert; Williams, Luke T .; Williams, Tim M .; Bolstridge, Mark; Sessa, Ben; McGonigle, John; Sereno, Martin I .; Nichols, David; Hellyer, Peter J .; Hobden, Peter; Evans, John; Singh, Krish D .; Wise, Richard G .; Curran, H. Valerie; Feilding, Amanda; Nutt, David J. (26. dubna 2016). „Neurální koreláty zkušeností s LSD odhalené multimodálním neuroimagingem“. Sborník Národní akademie věd. 113 (17): 4853–4858. Bibcode:2016PNAS..113.4853C. doi:10.1073 / pnas.1518377113. PMC 4855588. PMID 27071089.
- ^ Kringelbach, Morten L .; Green, Alexander L .; Aziz, Tipu Z. (2011-05-02). „Balancing the Brain: Resting State Networks and Deep Brain Stimulation“. Frontiers in Integrative Neuroscience. 5: 8. doi:10.3389 / fnint.2011.00008. PMC 3088866. PMID 21577250.
- ^ Sripada, Rebecca K .; King, Anthony P .; Welsh, Robert C .; Garfinkel, Sarah N .; Wang, Xin; Sripada, Chandra S .; Liberzon, Izrael (2012). „Neurální dysregulace u posttraumatické stresové poruchy“. Psychosomatická medicína. 74 (9): 904–911. doi:10.1097 / PSY.0b013e318273bf33. ISSN 0033-3174. PMC 3498527. PMID 23115342.
- ^ A b Akiki, Teddy J .; Averill, Christopher L .; Abdallah, Chadi G. (2017). „Síťově založený neurobiologický model PTSD: důkazy ze strukturálních a funkčních neuroimagingových studií“. Aktuální zprávy z psychiatrie. 19 (11): 81. doi:10.1007 / s11920-017-0840-4. ISSN 1523-3812. PMC 5960989. PMID 28924828.
- ^ Voss, Michelle W .; Soto, Carmen; Yoo, Seungwoo; Sodoma, Matthew; Vivar, Carmen; van Praag, Henriette (duben 2019). „Cvičení a hipokampální paměťové systémy“. Trendy v kognitivních vědách. 23 (4): 318–333. doi:10.1016 / j.tics.2019.01.006. PMC 6422697. PMID 30777641.
- ^ Shao, Mengling; Lin, Huiyan; Yin, Desheng; Li, Yongjie; Wang, Yifan; Ma, Junpeng; Yin, Jianzhong; Jin, Hua (2019-10-01). Rao, Hengyi (ed.). "Learning to play badminton altered resting-state activity and functional connectivity of the cerebellar sub-regions in adults". PLOS ONE. 14 (10): e0223234. doi:10.1371/journal.pone.0223234. ISSN 1932-6203. PMC 6771995. PMID 31574108.
- ^ Muraskin, Jordan; Dodhia, Sonam; Lieberman, Gregory; Garcia, Javier O.; Verstynen, Timothy; Vettel, Jean M .; Sherwin, Jason; Sajda, Paul (December 2016). "Brain dynamics of post-task resting state are influenced by expertise: Insights from baseball players: Brain Dynamics of Post-Task Resting State". Mapování lidského mozku. 37 (12): 4454–4471. doi:10.1002/hbm.23321. PMC 5113676. PMID 27448098.
- ^ Foster, Brett L .; Parvizi, Josef (2012-03-01). "Resting oscillations and cross-frequency coupling in the human posteromedial cortex". NeuroImage. 60 (1): 384–391. doi:10.1016/j.neuroimage.2011.12.019. ISSN 1095-9572. PMC 3596417. PMID 22227048.
- ^ Morris, Peter G.; Smith, Stephen M .; Barnes, Gareth R.; Stephenson, Mary C.; Hale, Joanne R.; Price, Darren; Luckhoo, Henry; Woolrich, Mark; Brookes, Matthew J. (2011-10-04). "Investigating the electrophysiological basis of resting state networks using magnetoencephalography". Sborník Národní akademie věd. 108 (40): 16783–16788. Bibcode:2011PNAS..10816783B. doi:10.1073/pnas.1112685108. ISSN 0027-8424. PMC 3189080. PMID 21930901.
- ^ Fair, D. A.; Cohen, A. L.; Dosenbach, N. U. F.; Church, J. A .; Miezin, F. M.; Barch, D. M.; Raichle, M. E.; Petersen, S. E.; Schlaggar, B. L. (2008). "The maturing architecture of the brain's default network". Sborník Národní akademie věd. 105 (10): 4028–32. Bibcode:2008PNAS..105.4028F. doi:10.1073/pnas.0800376105. PMC 2268790. PMID 18322013.
- ^ Morcom, Alexa M.; Fletcher, Paul C. (October 2007). "Does the brain have a baseline? Why we should be resisting a rest". NeuroImage. 37 (4): 1073–1082. doi:10.1016/j.neuroimage.2006.09.013. PMID 17052921.