Šupinatý šnek - Scaly-foot snail
| Šnek šupinatý / mořský luskoun | |
|---|---|
 | |
| Chrysomallon squamiferum od Longqi. Měřítko je 1 cm. | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Měkkýš |
| Třída: | Gastropoda |
| Clade: | Neomphalina |
| Rodina: | Peltospiridae |
| Rod: | Chrysomallon Chen, Linse, Copley & Rogers, 2015 |
| Druh: | C. squamiferum |
| Binomické jméno | |
| Chrysomallon squamiferum Chen, Linse, Copley & Rogers, 2015[2] | |
| Synonyma[2] | |
Crysomallon squamiferum (orth. chyba) | |

Chrysomallon squamiferum, běžně známý jako the šupinatá stopa, šupinatý šneknebo mořský pangolin[3], je druh hlubinných hydrotermální ventilace hlemýžď, a námořní plži měkkýš v rodině Peltospiridae.[2] Tento plicní endemický plže je známý pouze z hlubinných hydrotermálních průduchů v Indický oceán, kde byla nalezena v hloubkách asi 2 400–2 900 m (1,5–1,8 mil). Chrysomallon squamiferum se výrazně liší od ostatních hlubinných plžů, dokonce i od příbuzných neomfalinů.[4] V roce 2019 to bylo deklarováno ohrožený na Červeném seznamu IUCN[5], první druh, který má být uveden jako takový kvůli rizikům z hlubinná těžba jeho větracího stanoviště, které také produkuje vysoce kvalitní kovové rudy[6].
Plášť má jedinečnou konstrukci se třemi vrstvami; vnější vrstva se skládá z sulfidy železa, střední vrstva je ekvivalentní organické periostracum nachází se v jiných plžech a nejvnitřnější vrstva je vyrobena z aragonit. Noha je také neobvyklá a po stranách je pancéřována železem mineralizovanými sklerity.
Šnek jícnu domy symbiotický gamaproteobakterie ze kterého se zdá, že hlemýžď získává svou výživu. Tento druh je považován za jednoho z nejpodivnějších hlubinných hydrotermálních plžů a je jediným známým existujícím živočichem, který do své kostry (jak do skleritů, tak do skořápky jako exoskeleton) vnáší sulfid železa.[2] Jeho srdce je proporcionálně neobvykle velké pro jakékoli zvíře: srdce tvoří přibližně 4% jeho tělesného objemu.[4]
Taxonomie
Tento druh byl poprvé objeven v dubnu 2001 a od roku 2001 je označován jako „šupinatý“ plži.[7] Byl označován jako Chrysomallon squamiferum od roku 2003, ale nebyl formálně popsán ve smyslu Mezinárodní kodex zoologické nomenklatury až do Chen et al. pojmenoval to v roce 2015.[2][8] Typové vzorky jsou uloženy v Natural History Museum, Londýn.[2] V době, kdy název ještě nebyl formován, byla nesprávná pravopisná varianta „Crysomallon squamiferum".[2]
C. squamiferum je druh druhu a jediný druh rodu Chrysomallon.[2] Obecné jméno Chrysomallon je z Starořečtina jazyk a znamená „zlaté vlasy“, protože pyrit (sloučenina vyskytující se v její skořápce) má zlatou barvu.[2] Konkrétní název squamiferum pochází z latinského jazyka a znamená „šupinatý“, protože obsahuje sklerity.[2] Zpočátku nebylo známo, do které rodiny tento druh patří.[7] Warén et al. zařadil tento druh do rodiny Peltospiridae v rámci skupiny Neomphalina v roce 2003.[9] Molekulární analýzy založené na sekvencích cytochrom-c oxidáza I (COI) geny potvrdilo umístění tohoto druhu v Peltospiridae.[2][10] Morfotypy ze dvou lokalit jsou tmavé; morfototyp ze třetí lokality je bílý (vysvětlení lokalit viz další část).[2][11][12] Podle výsledků genetické analýzy se tyto různě zbarvené šneky zdají být jednoduše „odrůdami“ stejného druhu.[2]
Rozdělení

Šupinatý škeble je gastropod s větracími otvory, známý pouze z hlubinných hydrotermálních průduchů Indického oceánu, které jsou hluboké přibližně 2780 metrů.[2] Tento druh byl objeven v roce 2001 a žil na základnách černí kuřáci v Kairei hydrotermální ventilace pole, 25 ° 19,239 's 70 ° 02,429 'východní délky / 25,320650 ° j. 70.040483 ° v, na středoindickém hřebeni, severně od Rodrigues Triple Point.[7] Tento druh byl následně také nalezen na poli solitairu, 19 ° 33,413 's 65 ° 50,888 'východní délky / 19,556883 ° J 65,848133 ° E, Central Indian Ridge, v rámci Výhradní ekonomická zóna z Mauricius[13][14] a Longqi (v čínštině znamená „dračí vlajka“)[15] pole, 37 ° 47,027 's 49 ° 38,963 'východní délky / 37,783783 ° S 49,649383 ° E, Jihozápadní indický hřeben.[16][17] Pole Longqi bylo označeno jako typová lokalita; veškerý typový materiál pochází z tohoto odvětrávacího pole.[2] Vzdálenost mezi Kairei a Solitaire je asi 700 km (430 mi). Vzdálenost mezi Solitaire a Longqi je asi 2500 km (1600 mi).[2] Tyto tři lokality patří do Indického oceánu biogeografická provincie hydrotermálních ventilačních systémů Sensu Rogersi et al. (2012).[18] Vzdálenost mezi místy je velká, ale celková distribuční plocha je velmi malá, méně než 0,02 čtverečních kilometrů (0,0077 čtverečních mil).[19]
Je známo, že hlemýždi Peltospiridae žijí hlavně ve větrných polích východního Pacifiku. Nakamura et al. předpokládal, že výskyt šupinatých plžů v Indickém oceánu naznačuje vztah hydrotermálních ventilačních faun mezi těmito dvěma oblastmi.[13]Výzkumné expedice zahrnovaly:
- 2000 - expedice Japonská agentura pro vědu a technologii mořské Země pomocí lodi RV Kairei a ROV Kaiko objevil odvzdušňovací pole Kairei, ale v té době nebyli nalezeni šupinatí škeble.[20] Toto bylo první větrací pole objevené v Indickém oceánu.[20]
- 2001 - expedice amerického výzkumného plavidla RV Knorr s ROV Jasone objevili šupinaté plže ve větrném poli Kairei.[7]
- 2007 - expedice RV Da Yang Yi Hao objevili ventilační pole Longqi.[2]
- 2009 - expedice RV Yokosuka s DSV Shinkai 6500 objevili pole Solitaire a odebrali tam plže šupinatých stop.[13]
- 2009 - expedice RV Da Yang Yi Hao vizuálně pozorovaní šupinatí škeble na plicním poli Longqi.[2][16]
- 2011 - expedice britské královské výzkumné lodi RRS James Cook s ROV Kiel 6000 ochutnal ventilační pole Longqi.[2][21]
Popis
Sclerites

U tohoto druhu jsou boky šnečí nohy extrémně neobvyklé v tom, že jsou obrněné stovkami železem mineralizovaných sklerity; jsou složeny ze sulfidů železa[9] greigit a pyrit.[22] Každý sklerit má měkký epiteliální tkáňové jádro, a konchiolin kryt a nejvyšší vrstva obsahující pyrit a greigit.[2] Před objevením šupinatého plže se předpokládalo, že ve třídách jsou jediní existující měkkýši, kteří mají struktury podobné šupinám. Caudofoveata, Solenogastres a Polyplacophora.[17] Scleriti nejsou homologní k plži operculum. Skleriti šupinatých plžů také nejsou homologní se sklerity nalezenými v chitony (Polyplacophora).[17] Předpokládala se hypotéza, že skleriti kambrijští halwaxiids jako Halkieria mohou být potenciálně více analogické skleritům tohoto šneka, než jsou sklerity chitonů nebo aplakoforanů.[17] Ještě v roce 2015 nebyla provedena podrobná morfologická analýza pro testování této hypotézy.[17]
Sklerité z Chrysomallon squamiferum jsou převážně bílkovinné (conchiolin je komplexní protein); na rozdíl od toho jsou sklerity chitonů převážně vápenaté.[17] V příčných řezech skleritů nejsou viditelné žádné růstové linie konchiolinu.[17] Žádní jiní existující nebo vyhynulí plži nemají dermální sklerity,[17] a není známo žádné jiné existující zvíře, které by tímto způsobem používalo sulfidy železa kostra,[2] nebo exoskeleton.
Velikost každého skleritu je u dospělých asi 1 × 5 mm.[2] Mladiství mají váhy v několika řadách, zatímco dospělí mají husté a asymetrické váhy.[23] Populace solitairů hlemýžďů má místo černých bílé sklerity; je to způsobeno nedostatkem železa ve skleritech.[17] Skleriti jsou nasyceni (překrývají se způsobem připomínajícím střešní tašky ).[4] Spekuluje se, že účelem skleritů je ochrana nebo detoxikace.[24] Sklerity mohou pomoci chránit plži před ventilační tekutinou, takže její bakterie mohou žít v blízkosti zdroje dárců elektronů pro chemosyntézu.[4] Alternativně mohou sklerity vznikat usazováním toxických látek sulfid odpad z endosymbiontů, a proto představují nové řešení pro detoxikaci.[4] Skutečná funkce skleritů však zatím není známa.[13] Sklerity populace Kairei, které mají vrstvu sulfidu železa, jsou ferimagnetický.[2] Slerit mineralizovaný neželezným železem z morfotypu Solitaire se ukázal vyšší mechanická síla celé konstrukce v zátěžovém testu tříbodovým ohybem (12,06 MPa) než sklerit z morfotypu Kairei (6,54 MPa).[13]
V životě jsou na vnějším povrchu skleritů různé druhy epibionty: Epsilonproteobakterie a Deltaproteobakterie.[25] Tyto bakterie pravděpodobně poskytují svou mineralizaci.[25] Goffredi et al. (2004) předpokládali, že hlemýžď vylučuje některé organické sloučeniny, které usnadňují připojení bakterií.[25]
Shell

Kairei, Longqi, Solitaire (zleva doprava)
Skořápka těchto druhů má tři přesleny.[2] Tvar pláště je kulovitý a věž je komprimován.[2] Socha skořápky se skládá z žeber a jemných linií růstu.[2] Tvar clona je eliptický.[2] The vrchol skořápka je křehká a u dospělých koroduje.[2]
Jedná se o velmi velkou peltospiridu ve srovnání s většinou ostatních druhů, které jsou obvykle pod 15 milimetrů (3⁄5 v) v délce skořápky.[2] Šířka pláště je 9,80–40,02 mm (0,39–1,58 palce);[2] maximální šířka pláště dosahuje 45,5 milimetrů (1,79 palce).[2] Průměrná šířka skořápky dospělých hlemýžďů je 32 mm.[2] Průměrná šířka skořápky v populaci Solitaire byla o něco menší než v populaci Kairei.[14] Výška pláště je 7,65–30,87 mm (0,30–1,22 palce).[2] Šířka otvoru je 7,26–32,52 mm (0,29–1,28 palce).[2] Výška clony je 6,38–27,29 mm (0,25–1,07 palce).[2]
Šnečí ulita je také neobvyklá. Struktura pláště se skládá ze tří vrstev. Vnější vrstva je asi 30 μm silná, černá a je vyrobena ze sulfidů železa, které obsahují greigit Fe3S4.[26] Díky této vlastnosti je tento plže jediným dosud známým dosud existujícím zvířetem, které tento materiál ve své kostře využívá.[2] Střední vrstva (asi 150 μm) je ekvivalentní organickému periostracum, které se také vyskytuje u jiných plžů.[26] Periostracum je silné a hnědé.[2] Nejvnitřnější vrstva je vyrobena z aragonit (asi 250 μm silná), forma uhličitan vápenatý který se běžně vyskytuje jak ve skořápkách měkkýšů, tak v různých korálech.[26] Barva aragonitové vrstvy je mléčně bílá.[2]
Zdá se, že každá vrstva skořápky přispívá k účinnosti obrany hlemýžďů různými způsoby. Zdá se, že střední organická vrstva absorbuje mechanické namáhání a energii generovanou mačkáním (například drápy kraba), což činí skořápku mnohem tvrdší. Organická vrstva také působí na odvod tepla.[27] Vlastnosti tohoto Kompozitní materiál se zaměřují na výzkumné pracovníky pro možné použití v civilních a vojenských ochranných aplikacích.[26]
 Chrysomallon squamiferum z odvzdušňovacího pole Kairei. |  C. squamiferum z větracího pole Solitaire. |
Operculum
U tohoto druhu je tvar operculum změny během růstu, od zaobleného tvaru u mladistvých po zakřivený tvar u dospělých.[13] Relativní velikost opercula klesá s růstem jednotlivců.[4] Asi polovina všech dospělých hlemýžďů tohoto druhu má operculum mezi sklerity v zadní části zvířete.[13] Zdá se pravděpodobné, že sklerity postupně rostou a plně chrání celou nohu kvůli ochraně a operculum ztrácí svou ochrannou funkci, jak zvíře roste.[13]
 Mladistvý s operculum označený červeným ukazatelem. Délka pláště je asi 2 mm. |  Operculum mladistvého šneka. Měřítko je 1 mm. |  Operculum dospělého šneka. Měřítko je 1 mm. |  Dospělí šneci s operculum označeny červenými šipkami. Měřítko je 5 mm. |
Vnější anatomie
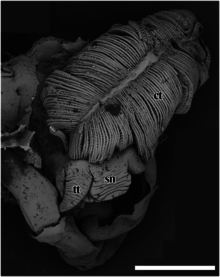
Šupinatá stopa má velký čenich, který se distálně zužuje na tupý konec. Ústa jsou kruhový kruh svalů, když jsou uzavřeny a uzavřeny.[4] Dva hladké cefalické chapadla jsou silné u základny a postupně se zužují do jemného bodu na jejich distálních koncích.[4] Tento šnek nemá oči.[4] Neexistuje žádný specializovaný kopulační dodatek.[4] Noha je červená a velká a hlemýžď ji nemůže úplně stáhnout do skořápky.[2] Tady není žádný pedální žláza v přední části chodidla.[4] Neexistují také žádná epipodiální chapadla.[4]
Vnitřní anatomie
v Chrysomallon squamiferum, měkké části zvířete zabírají přibližně dva kroužky vnitřku skořápky.[4] Skořepina je ve tvaru podkovy a velká, rozdělená na dvě části vlevo a vpravo a spojená užším připojením.[4] Okraj pláště je silný, ale jednoduchý bez výrazných rysů.[4] The dutina pláště je hluboká a dosahuje k zadnímu okraji skořápky.[4] Mediální levé straně dutiny dominuje velmi velký bipektinát ctenidium.[4] Ventrálně k viscerální hmotě je tělesná dutina obsazena obrovskou jícnovou žlázou, která se rozprostírá a vyplňuje ventrální dno dutiny pláště.[4][25]
The zažívací ústrojí je jednoduchý a je snížen na méně než 10% objemu typického pro plži.[4][25] The radula je „slabý“ rhipidoglosanového typu s jediným párem radikálních chrupavek.[4][25] The vzorec radule je ∼50 + 4 + 1 + 4 + ∼50.[2] Radula stuha je 4 mm dlouhá, 0,5 mm široká;[2] poměr šířky k délce je přibližně 1:10.[4] Tady není žádný čelist a žádné slinné žlázy.[4] Část předního jícnu rychle expanduje do obrovské, hypertrofované slepé jícnové žlázy se slepým koncem, která zaujímá velkou část ventrální strany dutiny pláště (odhaduje se na 9,3% objemu těla).[4] Když hlemýžď roste, jícnová žláza se zvyšuje izometricky s růstem.[23] Ezofageální žláza má jednotnou strukturu a je vysoce vaskularizovaná jemnými cévami.[4] The žaludek má v pravé přední části alespoň tři kanály, které se připojují k zažívací žláza.[4] V žaludku i v zadním střevě jsou konsolidované pelety.[4] Tyto pelety jsou pravděpodobně granule síra produkovaný endosymbiontem jako způsob detoxikace sirovodíku.[4] Střevo je zmenšené a má pouze jednu smyčku.[4] Rozsáhlá a nekonsolidovaná zažívací žláza sahá až k zadní části a vyplňuje skořápku vrchol skořápky.[4] Rektum neproniká do srdce, ale prochází ventrálně.[4] The řiť se nachází na pravé straně šneka, nad genitálním otvorem.[4]
V vylučovací systém, nephridium je centrální a směřuje k pravé straně těla jako tenká tmavá vrstva žlázové tkáně.[4] Nephridium je přední a ventrální zažívací žlázy a je v kontaktu s hřbetní stranou předního střeva.[4]
The dýchací systém a oběhový systém se skládají z jediného levého bipektinátu ctenidium (žába), která je velmi velká (15,5% objemu těla) a je podporována mnoha velkými a pohyblivými krevními dutinami naplněnými hemocoel.[4][23] Při pitvě jsou v dutině těla prominentní rysy krevní dutiny a hrudky materiálu hemocoel.[4] Ačkoli oběhový systém v Chrysomallon je většinou uzavřený (což znamená, že hemocoel většinou nezanechává krevní dutiny), prominentní krevní dutiny se zdají být přechodné a vyskytují se v různých částech těla u různých jedinců.[23] Na obou stranách ctenidia jsou tenká žaberní vlákna.[4] Bipektinát ctenidium sahá daleko za srdce do přeslenů horní skořápky; je mnohem větší než v Peltospira. Ačkoli tento druh má podobný tvar skořápky a obecnou formu jako jiné peltospiridy, ctenidium je úměrné velikosti k velikosti Hirtopelta, který má dosud největší žábru mezi rody peltospiridů, které byly dosud anatomicky zkoumány.[4]
Ctenidium poskytuje šneku kyslík, ale oběhový systém je zvětšen nad rámec jiných podobných plicních plžů.[4] V žábru nebo na žábu nejsou žádné endosymbionty C. squamiferum.[4] Zvětšení žábry má pravděpodobně usnadnit extrakci kyslíku v podmínkách s nízkým obsahem kyslíku, které jsou typické pro hydrotermální ventilační ekosystémy.[4]
Na zadní ctenidia je pozoruhodně velké a dobře vyvinuté srdce.[4] Srdce je pro každé zvíře proporcionálně neobvykle velké.[4] Na základě objemu jediného ušního boltce a komory představuje srdeční komplex přibližně 4% objemu těla (například srdce člověka je 1,3% objemu těla).[4] Komora má u mladistvého zvířete velikost 0,64 mm s délkou skořápky 2,2 mm a u dospělých dosáhne velikosti 8 mm.[23] Toto proporčně obří srdce primárně saje krev přes ctenidium a dodává vysoce vaskularizovanou jícnovou žlázu.[4] v C. squamiferum endosymbionty jsou umístěny v jícnu, kde jsou izolovány z ventilační tekutiny.[4] Je tedy pravděpodobné, že hostitel bude hrát hlavní roli při zásobování endosymbiontů potřebnými chemikáliemi, což povede ke zvýšeným respiračním potřebám.[4] Podrobné vyšetřování hemocoelu z C. squamiferum odhalí další informace o jeho respiračních pigmentech.[4]

ct - ctenidium,
pm - pedálový sval,
sc - váhy,
si - krevní sinus,
te - varle.
Měřítko je 1 cm.
Šupinatý škeble je chemosymbiotikum holobiont.[25] Hostuje thioautotrofní (síry oxidující) gamaproteobakteriální endosymbionty v mnohem větším jícnu, a zdá se, že spoléhá na tyto symbionty pro výživu.[25][28] Nejbližší známý příbuzný tohoto endosymbiontu je ten z Alviniconcha šneci.[29] U tohoto druhu je velikost jícnové žlázy asi o dva řády větší než obvyklá velikost.[25] V pažerákové žláze je významné zúžení, kde krevní tlak pravděpodobně klesá téměř na nulu.[4] Propracovaný kardiovaskulární systém se s největší pravděpodobností vyvinul tak, aby okysličoval endosymbionty v prostředí chudém na kyslík a / nebo aby dodával sirovodík na endosymbionty.[4] Thioautotrophic gammaproteobacteria have a full set of gens required for aerobní dýchání a jsou pravděpodobně schopné přepínat mezi efektivnějším aerobním dýcháním a méně účinným anaerobním dýcháním v závislosti na dostupnosti kyslíku.[4] V roce 2014 se endosymbiont šupinatého plže stal prvním endosymbiontem jakéhokoli plže, pro který byl znám celý genom.[28] C. squamiferum byl dříve považován za jediný druh Peltospiridae, který má zvětšenou jícnovou žlázu,[2] ale později se zjistilo, že oba druhy Gigantopelta mají také zvětšenou jícnovou žlázu.[10] Chrysomallon a Gigantopelta jsou jedinými větracími zvířaty, kromě siboglinid tubusové červy, které obsahují endosymbionty v uzavřené části těla, které nejsou v přímém kontaktu s ventilační tekutinou.[23]
The nervový systém je velký a mozek je pevná nervová hmota bez ganglií.[4] Nervový systém je ve srovnání s jinými taxony neomphaline složitější a větší.[4] Jak je typické pro plže, nervový systém se skládá z předního jícnového nervového prstence a dvou párů podélných nervových šňůr, přičemž ventrální pár inervuje nohu a hřbetní pár tvořící zákrut streptoneury.[4] Čelní část jícnového nervového prstence je velká a spojuje dva boční otoky.[4] Obrovská fúzovaná nervová hmota přímo sousedí s eosofageální žlázou a prochází jícnem, kde jsou umístěny bakterie.[4] Do hlavových chapadel vyčnívají velké chapadlové nervy.[4] The smyslové orgány šupinatého plže patří statocysty obklopena jícnovou žlázou, každá statocystou jedinou statolit.[4] Na špičce žaberních vláken jsou také senzorické ctenidiální bursicles; tito jsou známí být přítomný ve většině vetigastropods, a být přítomný některý neomphalines.[4]
The rozmnožovací systém má některé neobvyklé vlastnosti. Pohlavní žlázy dospělých hlemýžďů nejsou uvnitř pláště; jsou v oblasti hlavy a nohou na pravé straně těla.[4] U mladistvých s délkou pláště 2,2 mm nejsou přítomny žádné pohlavní žlázy.[23] Dospělí mají obojí varle a vaječník na různých úrovních vývoje.[4] Varle je umístěno ventrálně; vaječník je uložen dorzálně a nefridium leží mezi nimi.[4] Vedle varlat je „obalový orgán spermatoforů“.[4] Gonodukty ze varlete a vaječníku jsou zpočátku oddělené, ale zjevně se spojují do jediného vývodu a vystupují jako jediný genitální otvor napravo od dutiny pláště.[4] Zvíře nemá kopulační orgán.[2][4]
 Hřbetní pohled na Chrysomallon squamiferum zobrazující přehled dutiny pláště. Plášť a tkáň pláště byly odstraněny. Měřítko je 1 cm. |  3D rekonstrukce ukazuje velké ctenidium a velké srdce, hřbetní pohled. Měřítko je 250 μm.
|  3D rekonstrukce trávicího systému ukazuje zvětšenou jícnovou žlázu, hřbetní pohled. Měřítko je 250 μm.
|
Předpokládá se, že odvozená strategie umístění endosymbiotických mikrobů v jícnové žláze byla katalyzátorem anatomických inovací, které slouží především ke zlepšení kondice bakterií, nad rámec potřeb šneka.[4] Velké zvětšení jícnové žlázy, hlemýžďovy ochranné dermální sklerity, jeho vysoce rozšířený dýchací a oběhový systém a jeho vysoká plodnost jsou považovány za adaptace, které jsou prospěšné pro jeho endosymbiontové mikroby.[4] Tyto úpravy se zdají být výsledkem specializace na extrémní řešení energetických potřeb chemosyntetický životní prostředí.[4]
Ekologie
Místo výskytu

Tento druh obývá hydrotermální větrací pole Indického oceánu. Žije v sousedství jak kyselé, tak redukční ventilační tekutiny, na stěnách černých kuřáckých komínů nebo přímo na difúzních místech proudění.[4]
Hloubka pole Kairei se pohybuje od 2 415 do 2 460 m (7 923 až 8 071 stop),[7] a jeho rozměry jsou přibližně 30 x 80 m (98 x 262 ft).[7] Sklon pole je 10 ° až 30 °.[7] The Podklad rock je troctolit a vyčerpaný středooceánský hřeben čedič.[30] Kairei-polní šupinatí plži žijí v nízkoteplotních difúzních tekutinách jediného komína.[13] Přechodná zóna, kde byli tito plži nalezeni, má šířku asi 1–2 m (3–7 ft) a teplotu 2–10 ° C.[31] Preferovaná teplota vody pro tento druh je asi 5 ° C.[32] Tito šneci žijí v prostředí, které má vysoké koncentrace sirovodíku a nízké koncentrace kyslíku.[32]
Množství šupinatých plžů bylo v poli Kairei nižší než v poli Longqi.[2] Komunita hydrotermálních větrání Kairei se skládá z 35 taxonů,[33] včetně mořských sasanek Marianactis sp., korýši Austinograea rodriguezensis, Rimicaris kairei, Mirocaris indica, Munidopsis sp., Neolepadidae rod a sp., Eochionelasmus sp., mlži Bathymodiolus marisindicus, plži Lepetodrilus sp., Pseudorimula sp., Eulepetopsis sp., Shinkailepas sp. a Alviniconcha marisindica,[34] Desbruyeresia marisindica,[35] Bruceiella wareni,[35] Phymorhynchus sp., Sutilizona sp., slit limpet sp. 1, štěrbina limpet sp. 2, Iphinopsis boucheti,[35] solenogastres Helicoradomenia ? sp., annelids Amphisamytha sp., Archinome jasoni, Capitellidae sp. 1, Ophyotrocha sp., Hesionoidae sp. 1, Hesionoidae sp. 2, Branchinotogluma sp., Branchipolynoe sp., Harmothoe ? sp., Levensteiniella ? sp., Prionospio sp., neznámý Nemertea a neidentifikovaný Platyhelminthes.[33] Šupinatá stopa plži žijí v koloniích s Alviniconcha marisindica šneci, a tam jsou kolonie Rimicaris kairei nad nimi.[32]
Pole solitairu je v hloubce 2 606 m (8 550 ft) a jeho rozměry jsou přibližně 50 x 50 m (160 x 160 ft).[13] Hornina substrátu je obohacený středooceánský hřebenový čedič.[13][30] Šupinatí plži žijí poblíž vysokoteplotních difúzních tekutin komínů ve větracím poli.[13] Množství šupinatých plžů bylo nižší než v poli Longqi.[2] Solitaire hydrotermální ventilační komunita zahrnuje 22 taxonů, včetně: mořských sasanek Marianactis sp., korýši Austinograea rodriguezensis, Rimicaris kairei, Mirocaris indica, Munidopsis sp., Neolepadidae gen et sp., Eochionelasmus sp., mlži Bathymodiolus marisindicus, plži Lepetodrilus sp., Eulepetopsis sp., Shinkailepas sp., Alviniconcha sp. typ 3, Desbruyeresia sp., Phymorhynchus sp., annelids Alvinellidae rod a sp., Archinome jasoni, Branchinotogluma sp., ostnokožci holothurians Apodacea gen et sp., ryby Macrouridae rod a sp., neznámý Nemertea a neidentifikovaný Platyhelminthes.[33]

Větrací pole Longqi je v hloubce 2 780 m (9 120 ft),[2] a jeho rozměry jsou přibližně 100 x 150 m (330 x 490 ft).[19] Chrysomallon squamiferum byla hustě osídlena v oblastech bezprostředně obklopujících ventilaci s difuzním tokem.[4] Komunita hydrotermálních ventilací Longqi zahrnuje 23[Poznámka 1] taxony makro- a megafauna: sasanky Actinostolidae sp., annelids Polynoidae n. gen. n. sp. „655“, Branchipolynoe n. sp. "Drak", Peinaleopolynoe n. sp. "Drak", Hesiolyra srov. bergi, Hesionidae sp. indet., Ophryotrocha n. sp. „F-038 / 1b“, Prionospio srov. unilamellata, Ampharetidae sp. indet., mušle Bathymodiolus marisindicus, plži Gigantopelta aegis,[10] Dracogyra subfuscus, Lirapex politus,[15] Phymorhynchus n. sp. „SWIR“, Lepetodrilus n. sp. „SWIR“, korýši Neolepas sp. 1, Rimicaris kairei, Mirocaris indica, Chorocaris sp., Kiwa n. sp. „SWIR“ 17, Munidopsis sp. a ostnokožců holothurians Chiridota sp.[16][36] Hustota Lepetodrilus n. sp. „SWIR“ a šupinatí škeble jsou více než 100 hlemýžďů na m² v těsné vzdálenosti od zdrojů ventilačních tekutin na větracím poli Longqi.[36]
Stravovací návyky
Šupinatý škeble je povinný symbiotrof po celý život po vypořádání.[23] Výživa šupinatého plže po celou dobu jeho post-larválního života závisí na chemoautotrofie jejích endosymbiotických bakterií, které zajišťují veškerou její výživu.[25][23] Škeble-noha gastropod není ani filtr krmítko[4][23] ani nepoužívá jiné mechanismy krmení.[4] Radula se skládá pouze z 0,4% objemu těla u mladistvých a radula chrupavky se skládá z 0,8% objemu těla u mladistvých,[23] protože se již nepoužívají ke krmení.
Pro identifikaci trofické interakce v prostředí, kde je přímé pozorování stravovacích návyků komplikované, byly měřeny stabilní izotopové složení uhlíku a dusíku.[31] Existují vyčerpané hodnoty δ13C v jícnu (relativně k fotosynteticky odvozenému organickému uhlíku).[25] Za zdroj takového uhlíku se považovaly chemoautotrofní symbionty.[25] Chemoautotrofní původ stabilního izotopu uhlíku 13C bylo potvrzeno experimentálně.[28]
| tkáň | δ13C | δ15N |
|---|---|---|
| jícnu | −20.7 ± 0.9 ‰ | 3.3 ± 1.8 ‰ |
| žábry | −18,3 ± 0,6 ‰, od −17,4 do −18,8 ‰ | 3,9 ± 0,6 ‰, od 3,1 do 4,2 ‰ |
| plášť | od −17,5 do −18,6 ‰ | od 3,5 do 4,7 ‰ |
| chodidlo | −18.2 ± 0.6 ‰ | 3.8 ± 0.5 ‰ |
| váhy | −16.7 ± 0.6 ‰ | 3.8 ± 0.9 ‰ |
Životní cyklus
Tento plže je a simultánní hermafrodit.[4] Je to jediný druh z čeledi Peltospiridae, o kterém je dosud známo, že je současně hermafroditem.[4] Má vysokou úrodnost.[4] Klade vajíčka, která jsou pravděpodobně lecitotrofního typu.[21] Vejce šupinatých plžů vykazují při atmosférickém tlaku negativní vztlak.[14] Ani larvy, ani protoconch je známý od roku 2016, ale předpokládá se, že tento druh má fázi šíření planktonu.[21] Nejmenší Chrysomallon squamiferum juvenilní vzorky, které kdy byly odebrány, měly délku skořápky 2,2 mm.[23] Výsledky statistických analýz neodhalily žádnou genetickou diferenciaci mezi dvěma populacemi v polích Kairei a Solitaire, což naznačuje možnou konektivitu mezi těmito dvěma ventilačními poli.[14] Populace Kairei představuje potenciální zdrojovou populaci pro dvě populace ve středoindickém hřebeni.[14] Tyto šneky je těžké udržet naživu v umělém prostředí, přežili však v akváriích za atmosférického tlaku déle než tři týdny.[32]
Ochranná opatření a hrozby
Šupinatá stopa není chráněna.[1][19] Jeho celková distribuční plocha se odhaduje na maximálně 0,27 km2, omezeno na tři hlášené lokality, mezi nimiž dochází pouze k zanedbatelné migraci.[37] Hydrotermální průduchy v jihozápadním Indian Ridge se šíří pomalu a jeho komunity jsou považovány za citlivější na rušení a s pomalým zotavováním.[19] U tohoto druhu existuje riziko možného poškození životního prostředí hlubinná těžba.[19] Komerční licence k průzkumu těžby do oblasti Kairei byla udělena Mezinárodní úřad pro mořské dno do Německa od roku 2015 do roku 2030.[19] Licence na komerční průzkum těžby do oblasti Longqi byla udělena Číně od roku 2011 do roku 2026.[19] Na seznamu byl uveden jako ohrožený druh Červený seznam ohrožených druhů IUCN dne 4. července 2019.[1]
Poznámky
- ^ Od roku 2016 bylo z Longqi známo 21 druhů a v roce 2017 byly popsány dva nové plži.
Reference
Tento článek včlení text Creative Commons (CC-BY-4.0) z odkazů[4][14][23] a text CC-BY-2.5 z reference[13]
- ^ A b C „Červený seznam ohrožených druhů IUCN“. https://www.iucnredlist.org/species/103636217/103636261. 2019. Citováno 2019-07-18. Externí odkaz v
| web =(Pomoc) - ^ A b C d E F G h i j k l m n Ó str q r s t u proti w X y z aa ab ac inzerát ae af ag ah ai aj ak al dopoledne an ao ap vod ar tak jako na Chen, Chong; Linse, Katrin; Copley, Jonathan T .; Rogers, Alex D. (2015). „„ Plže šupinatá “: nový rod a druh hydrotermálního plicního endemického plže (Neomphalina: Peltospiridae) z Indického oceánu“. Journal of Molluscan Studies. 81 (3): 322–334. doi:10.1093 / měkkýš / eyv013.
- ^ Sigwart, Julia D .; Chen, Chong; Thomas, Elin A .; Allcock, A. Louise; Böhm, Monika; Seddon, Mary (2019-07-22). „Červený seznam může chránit hlubinnou biologickou rozmanitost“. Ekologie a evoluce přírody. 3 (8): 1134. doi:10.1038 / s41559-019-0930-2. ISSN 2397-334X. PMID 31332328.
- ^ A b C d E F G h i j k l m n Ó str q r s t u proti w X y z aa ab ac inzerát ae af ag ah ai aj ak al dopoledne an ao ap vod ar tak jako na au av aw sekera ano az ba bb před naším letopočtem bd být bf bg bh bi bj bk bl bm bn bo bp bq br bs bt bu bv bw Chen, Chong; Copley, Jonathan T .; Linse, Katrin; Rogers, Alex D .; Sigwart, Julia D. (2015). „Srdce draka: 3D anatomická rekonstrukce„ šupinatého plže “(Mollusca: Gastropoda: Neomphalina) odhaluje jeho mimořádný oběhový systém.“. Hranice v zoologii. 12: 13. doi:10.1186 / s12983-015-0105-1. PMC 4470333. PMID 26085836.
- ^ „Červený seznam ohrožených druhů IUCN“. Červený seznam ohrožených druhů IUCN. Citováno 2019-07-28.
- ^ Lambert, Jonathan (2019-07-22). „Oceánský šnek je první zvíře, které je oficiálně ohroženo hlubinnou těžbou“. Příroda. 571 (7766): 455–456. doi:10.1038 / d41586-019-02231-1. PMID 31337912.
- ^ A b C d E F G Dover, Cindy L. Van; Humphris, S.E .; Fornari, D .; Cavanaugh, C. M .; Collier, R .; Goffredi, Shana K .; Hashimoto, J .; Lilley, M. D .; Reysenbach, A.L .; Shank, T. M .; Von Damm, K. L .; Banta, A .; Gallant, R. M .; Gotz, D .; Green, D .; Hall, J .; Harmer, T. L .; Hurtado, L. A .; Johnson, P .; McKiness, Z. P .; Meredith, C .; Olson, E .; Pan, I.L .; Turnipseed, M .; Won, Y .; Young, C. R. 3.; Vrijenhoek, R. C. (2001). „Biogeografie a ekologické prostředí hydrotermálních průduchů Indického oceánu“. Věda. 294 (5543): 818–23. Bibcode:2001Sci ... 294..818V. doi:10.1126 / science.1064574. PMID 11557843.
- ^ Bouchet, P. (2014). "Chrysomallon squamiferum". Světový registr mořských druhů. Citováno 2015-04-22.
- ^ A b Warén, Anders; Bengtson, Stefan; Goffredi, Shana K .; Dover, Cindy L. Van (2003). "Horký-plži plži s sulfidem železitým dermálními sklerity". Věda. 302 (5647): 1007. doi:10.1126 / science.1087696. PMID 14605361.
- ^ A b C Chen, Chong; Linse, Katrin; Roterman, Christopher N .; Copley, Jonathan T .; Rogers, Alex D. (2015). „Nový rod velkých hydrotermálních plicních endemických plžů (Neomphalina: Peltospiridae)“ (PDF). Zoologický žurnál Linneanské společnosti (Vložený rukopis). 175 (2): 319–335. doi:10.1111 / zoj.12279.
- ^ (v japonštině) (2010) „硫化 鉄 を 纏 わ な い 白 ス ケ ー リ ー フ ッ ト を 世界 で 初 め て 発 見 ~ イ ン 洋 に おお. Japonská agentura pro vědu a technologii mořské Země, Tokijská univerzita, Univerzita Kochi. (tisková zpráva). Citováno 2016-07-16.
- ^ "Nový šupinatý škeble nalezený v Indickém oceánu; objev bílého šupinatého škeble". Southern Fried Science. 1. července 2011. Citováno 2016-07-16.
- ^ A b C d E F G h i j k l m Nakamura, Kentaro; Watanabe, Hiromi; Miyazaki, Junichi; Takai, Ken; Kawagucci, Shinsuke; Noguchi, Takuro; Nemoto, Suguru; Watsuji, Tomo-o; Matsuzaki, Takuya; Shibuya, Takazo; Okamura, Kei; Mochizuki, Masashi; Orihashi, Yuji; Ura, Tamaki; Asada, Akira; Marie, Daniel; Koonjul, Meera; Singh, Manvendra; Beedessee, Girish; Bhikajee, Mitrasen; Tamaki, Kensaku; Schnur, Joel M. (2012). „Objev nové hydrotermální aktivity a chemosyntetické fauny na středoindickém hřebeni při 18 ° –20 ° J“. PLOS ONE. 7 (3): e32965. Bibcode:2012PLoSO ... 732965N. doi:10.1371 / journal.pone.0032965. PMC 3303786. PMID 22431990.
- ^ A b C d E F Beedessee, Girish; Watanabe, Hiromi; Ogura, Tomomi; Nemoto, Suguru; Yahagi, Takuya; Nakagawa, satoshi; Nakamura, Kentaro; Takai, Ken; Koonjul, Meera; Marie, Daniel E. P. (2013). „Vysoká konektivita zvířecích populací v hlubinných hydrotermálních větracích polích ve středoindickém hřebeni relevantní pro jejich geologické prostředí“. PLOS ONE. 8 (12): e81570. Bibcode:2013PLoSO ... 881570B. doi:10.1371 / journal.pone.0081570. PMC 3864839. PMID 24358117.
- ^ A b Chen, Chong; Zhou, Yadong; Wang, Chunsheng; Copley, Jonathan T. (2017). „Dva nové hlemýžďové hlemýžďové peltospiridové hlemýždi (Gastropoda: Neomphalina) z hydrotermálního pole Longqi, jihozápadní indický hřeben“. Frontiers in Marine Science. 4. doi:10.3389 / fmars.2017.00392. ISSN 2296-7745.
- ^ A b C Tao, Chunhui; Lin, Jian; Guo, Shiqin; Chen, Yongshun John; Wu, Šanghaj; Han, Xiqiu; Němec, Christopher R .; Yoerger, Dana R .; Zhou, Ning; Li, Huaiming; Su, Xin; Zhu, Jian (2012). DY115-19 (nohy 1–2) a DY115-20 (nohy 4–7) Vědecké párty. „První aktivní hydrotermální průduchy na ultra-pomalu se šířícím centru: Southwest Indian Ridge“. Geologie. 40 (1): 47–50. Bibcode:2012Geo .... 40 ... 47T. doi:10.1130 / G32389.1.
- ^ A b C d E F G h i Chen, Chong; Copley, Jonathan T .; Linse, Katrin; Rogers, Alex D .; Sigwart, Julia (2015). „Jak měkkýši dostali své váhy: konvergentní vývoj skleromomu měkkýšů“. Biologický žurnál společnosti Linnean. 114 (4): 949–954. doi:10.1111 / bij.12462.
- ^ Rogers, Alex D .; Tyler, Paul A .; Connelly, Douglas P .; Copley, Jon T .; James, Rachael; Larter, Robert D .; Linse, Katrin; Mills, Rachel A .; Garabato, Alfredo Naveira; Pancost, Richard D .; Pearce, David A .; Polunin, Nicholas V. C .; Němec, Christopher R .; Shank, Timothy; Boersch-Supan, Philipp H .; Alker, Belinda J .; Aquilina, Alfred; Bennett, Sarah A .; Clarke, Andrew; Dinley, Robert J. J .; Graham, Alastair G. C .; Green, Darryl R. H .; Hawkes, Jeffrey A .; Hepburn, Laura; Hilario, Ana; Huvenne, Veerle A. I .; Marsh, Leigh; Ramirez-Llodra, Eva; Reid, William D. K .; Roterman, Christopher N .; Sweeting, Christopher J .; Thatje, Sven; Zwirglmaier, Katrin (2012). „Objev nových hlubinných hydrotermálních ventilačních komunit v jižním oceánu a důsledky pro biogeografii“. PLOS Biology. 10 (1): –1001234. doi:10.1371 / journal.pbio.1001234. ISSN 1545-7885. PMC 3250512. PMID 22235194.
- ^ A b C d E F G Sigwart, Julia D .; Chen, Chong; Marsh, Leigh (2017). „Je těžba mořského dna pro měkkýše špatná?“. Nautilus. 131 (1): 43–49.
- ^ A b Hashimoto, červen; Ach, Suguru; Gamo, Toshitaka; Chiba, Hitoshi; Yamaguchi, Toshiyuki; Tsuchida, Shinji; Okudaira, Takamoto; Watabe, Hajime; Yamanaka, Toshiro; Kitazawa, Mitsuko (2001). „Objeveny první hydrotermální ventilační komunity z Indického oceánu“. Zoologická věda. 18 (5): 717–721. doi:10,2108 / zsj.18.717.
- ^ A b C Chen, Chong; Copley, Jonathan T .; Linse, Katrin; Rogers, Alex D. (2015). „Nízká konektivita mezi populacemi„ šupinatých plžů “(Mollusca: Peltospiridae) u hydrotermálních průduchů na jihozápadním indiánském hřebeni a středoindickém hřebeni.“ Organismy Rozmanitost a evoluce. 15 (4): 663–670. doi:10.1007 / s13127-015-0224-8.
- ^ Pickrell, John (07.11.2003). „Brnění pozlacené objeveno v hlubokém moři“. Zprávy z National Geographic. Citováno 2016-07-16.
- ^ A b C d E F G h i j k l m Chen, Chong; Uematsu, Katsuyuki; Linse, Katrin; Sigwart, Julia D. (2017). "Více způsoby než jedním: Rychlá konvergence u hydrotermálních průduchů ukázaná 3D anatomickou rekonstrukcí Gigantopelta (Mollusca: Neomphalina) ". BMC Evoluční biologie. 17 (1): 62. doi:10.1186 / s12862-017-0917-z. ISSN 1471-2148. PMC 5333402. PMID 28249568.
- ^ Suzuki, Yohey; Kopp, Robert E .; Koruge, Toshihiro; Suga, Akinobu; Takai, Ken; Tsuchida, Shinji; Ozaki, Noriaki; Endo, Kazuyoshi; Hashimoto, červen; Kato, Yasuhiro; Mizota, Chitoshi; Hirata, Takafumi; Chiba, Hitoshi; Nealson, Kenneth H .; Horikoshi, Koki; Kirschvink, Joseph L. (2006). „Tvorba skleritů v hydrotermálním„ šupinatém “gastropodu - možná kontrola biomineralizace sulfidu železa zvířetem“ (PDF). Dopisy o Zemi a planetách. 242 (1–2): 39–50. Bibcode:2006E & PSL.242 ... 39S. doi:10.1016 / j.epsl.2005.11.029.
- ^ A b C d E F G h i j k l m Goffredi, Shana K .; Warén, Anders; Orphan, Victoria J .; Dover, Cindy L. Van; Vrijenhoek, Robert C. (5. května 2004). „Nové formy strukturální integrace mezi mikroby a hydrotermální plži z Indického oceánu“. Aplikovaná a environmentální mikrobiologie. 70 (5): 3082–3090. doi:10.1128 / AEM.70.5.3082-3090.2004. PMC 404406. PMID 15128570.
- ^ A b C d Yao, Haimin; Dao, Ming; Imholt, Timothy; Huang, Jamie; Wheeler, Kevin; Bonilla, Alejandro; Suresh, Subra; Ortiz, Christine (2010). "Protection mechanisms of the iron-plated armor of a deep-sea hydrothermal vent gastropod". PNAS. 107 (3): 987–992. Bibcode:2010PNAS..107..987Y. doi:10.1073/pnas.0912988107. PMC 2808221. PMID 20133823.
- ^ "Snail's iron armour eyed by military". CBC News. 19. 1. 2010. Citováno 2016-07-16.
- ^ A b C Nakagawa, Satoshi; Shimamura, Shigeru; Takaki, Yoshihiro; Suzuki, Yohey; Murakami, Shun-ichi; Watanabe, Tamaki; Fujiyoshi, So; Mino, Sayaka; Sawabe, Tomoo; Maeda, Takahiro; Makita, Hiroko; Nemoto, Suguru; Nishimura, Shin-Ichiro; Watanabe, Hiromi; Watsuji, Tomo-o; Takai, Ken (2014). "Allying with armored snails: the complete genome of gammaproteobacterial endosymbiont". Časopis ISME. 8 (1): 40–51. doi:10.1038/ismej.2013.131. PMC 3869010. PMID 23924784.
- ^ Distel, Daniel L.; Altamia, Marvin A.; Lin, Zhenjian; Shipway, J. Reuben; Han, Andrew; Forteza, Imelda; Antemano, Rowena; Limbaco, Ma Gwen J. Peñaflor; Tebo, Alison G.; Dechavez, Rande; Albano, Julie; Rosenberg, Gary; Concepcion, Gisela P.; Schmidt, Eric W.; Haygood, Margo G. (2017-04-17). "Discovery of chemoautotrophic symbiosis in the giant shipworm Kuphus polythalamia (Bivalvia: Teredinidae) extends wooden-steps theory". Sborník Národní akademie věd. 114 (18): E3652–E3658. doi:10.1073/pnas.1620470114. ISSN 1091-6490. PMC 5422788. PMID 28416684. Citováno 2017-04-18.
- ^ A b Nakamura, Kentaro; Takai, Ken (2015). "Indian Ocean Hydrothermal Systems: Seafloor Hydrothermal Activities, Physical and Chemical Characteristics of Hydrothermal Fluids, and Vent-Associated Biological Communities". In Ishibashi J.-i.; et al. (eds.). Podzemní biosféra spojená s hydrotermálními systémy. Springer, Tokio. 147–161. doi:10.1007/978-4-431-54865-2_12. ISBN 9784431548645.
- ^ A b C Dover, Cindy Van (2002). "Trophic relationships among invertebrates at the Kairei hydrothermal vent field (Central Indian Ridge)". Mořská biologie. 141 (4): 761–772. doi:10.1007/s00227-002-0865-y.
- ^ A b C d "Extensive population of a "rare" scaly-foot gastropod discovered". Japonská agentura pro vědu a technologii mořské Země, Univerzita Hokkaido, Enoshima akvárium. 30. listopadu 2009. Citováno 2016-07-16.
- ^ A b C Watanabe, Hiromi; Beedessee, Girish (2015). "Vent Fauna on the Central Indian Ridge". In Ishibashi J.-i.; et al. (eds.). Podzemní biosféra spojená s hydrotermálními systémy. Springer, Tokio. pp. 205–212. doi:10.1007/978-4-431-54865-2_16. ISBN 9784431548645.
- ^ Johnson, Shannon B.; Warén, Anders; Tunnicliffe, Verena; Dover, Cindy Van; Wheat, C. Geoffrey; Schultz, Thomas F .; Vrijenhoek, Robert C. (2015-05-04). "Molekulární taxonomie a pojmenování pěti kryptických druhů Alviniconcha hlemýždi (Gastropoda: Abyssochrysoidea) z hydrotermálních průduchů “. Systematika a biologická rozmanitost. 13 (3): 278–295. doi:10.1080/14772000.2014.970673. ISSN 1477-2000.
- ^ A b C Okutani, Takashi; Hashimoto, červen; Sasaki, Takenori (2004). "New gastropod taxa from a hydrothermal vent (Kairei Field) in the central Indian Ocean" (PDF). Venuše. 63 (1–2): 1–10. Archivovány od originál (PDF) dne 04.10.2013.
- ^ A b Copley, J. T .; Marsh, L .; Glover, A. G .; Hühnerbach, V .; Nye, V. E .; Reid, W. D. K .; Sweeting, C. J .; Wigham, B. D .; Wiklund, H. (2016). „Ekologie a biogeografie megafauny a makrofauny na prvních známých hlubinných hydrotermálních průduchech na jihozápadním indiánském hřebeni, který se rozprostírá velmi pomalu“. Vědecké zprávy. 6: 39158. Bibcode:2016NatSR...639158C. doi:10.1038 / srep39158. ISSN 2045-2322. PMC 5155287. PMID 27966649.
- ^ Sigwart, Julia D. (Winter 2017). "Deep-sea conservation and the 'scaly-foot gastropod'" (PDF). Chapadlo. 25: 39–40.