Fytoglobin - Phytoglobin
Zdá se, že hlavní přispěvatel do tohoto článku má úzké spojení s jeho předmětem. (Dubna 2017) (Zjistěte, jak a kdy odstranit tuto zprávu šablony) |
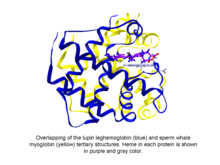
Fytoglobiny jsou kulovité rostlinné (řasy a suchozemské rostliny) bílkoviny zařazené do globin nadčeleď, která obsahuje hemu, i.E. protoporfyrin IX -Fe, protetická skupina. Nejjednodušší známé fytoglobiny jsou leghemoglobiny, objevený v roce 1939 Kubo po spektroskopické a chemické analýze červeného pigmentu sóji kořenové uzliny.[1] Několik desetiletí po Kuboově zprávě krystalizace fytoglobinu vlčího bobu (známého jako leghemoglobin) Vainshteinem a spolupracovníkem odhalila, že terciární struktura tohoto proteinu a spermie velryby myoglobin byl pozoruhodně podobný, což naznačuje, že fytoglobin objevený Kubo skutečně odpovídal globinu.[2]
Jednou z důležitých funkcí fytoglobinu je jeho oxid dusnatý dioxygenáza aktivita.[3]
Distribuce a klasifikace
Fytoglobiny (zkráceně Phytogbs) jsou všudypřítomně distribuovány v rostlinách, protože byly identifikovány v řasách a suchozemských rostlinách, včetně primitivních mechorosty a vyvinul se jednoděložní rostliny a dvouděložné rostliny. Mohou být klasifikovány následovně:[3][4]
- 2/2 fytoglobiny (zkrácená rodina globinu, podrodina TrHb2)
- Phytogb3: nachází se v řasách a suchozemských rostlinách.
- 3/3 fytoglobiny (rodina podobná myoglobinu)
- Phytogb0: bazální typ nacházející se v řasách, mechorosty a gymnospermy.[4]
- Phytogb1, 2: k dispozici pouze v krytosemenné rostliny.
Non-luštěniny symbiotické globiny (SymPhytogbs)[4] jsou rozptýleny mezi Phytogb1 a Phytogb2.[3] Symbiotické globiny obecně dodávají kyslík symbiotickým bakteriím, které provádějí fixaci dusíku. V luštěninách jsou bakterie rhizobia, ale v některých aktinorhizálních rostlinách aktinomycete Frankia místo toho udělejte práci.[3]
Strukturální charakteristiky
Fytogby jsou kódovány geny přerušenými 3 introny (i když obsahuje 4 introny fytogb gen byl detekován v mech Physcomitrella patens ).[5] První a třetí intron fytogb geny jsou lokalizovány ve stejné pozici jako geny myoglobin geny, což tomu nasvědčuje fytogb a myoglobin geny se vyvinuly ze společného předka před více než 600 miliony let.[6] Existence druhého intronu v fytogb geny předpověděl Go pomocí teoretické analýzy,[7] který byl dále ověřen klonováním a sekvenováním sója lb geny Markera a spolupracovníků.[8]

Fytogby jsou monomerní proteiny, jejichž molekulová hmotnost se pohybuje od ~ 17 do ~ 19 kDa. Při vysokých (více než 1 mM) koncentracích však mohou Phytogbs tvořit dimery. Polypeptidový řetězec fytogbs se skládá do zvláštního uspořádání 6 až 7 šroubovic (pojmenovaných písmeny A až H) známých jako globinový záhyb, který tvoří hydrofobní kapsu, kde je umístěn lem. Ve Phytogbs byly identifikovány dva typy globinového skládání: skládání 3/3 a 2/2,[9][10] kde se šroubovice A, E a F překrývají se šroubovicemi B, G a H a šroubovice B a E se překrývají se šroubovicemi G a H.

Stejně jako ostatní globiny je heme-Fe ve Phytogbs koordinován v proximální oblasti His aminokyselinou (pojmenovanou jako proximální His). Distální oblast hemu-Fe může být obsazena různými ligandy (např kyslík a oxid dusnatý ) nebo distální (často His) aminokyselina, která dává místo penta- nebo hexa-koordinaci Phytogbs. Phytogbs2, SymPhytogbs a Lbs jsou převážně penta-souřadnice, zatímco Phytogbs1 jsou převážně hexa-souřadnice a Phytogbs0 a Phytogbs3 jsou kombinací penta- a hexa-souřadnic.[4] Koordinace Heme-Fe je nezbytná pro funkci Phytogb (a dalších globinů), protože reguluje rychlost vazby a uvolňování ligandu v důsledku kinetických konstant kna a kvypnuto, resp. Například afinita sóji Lb a rýže Phytogb1 k O2 (K.Ó2) je mírný a velmi vysoký, protože kna je 130 a 68 mM−1 s−1, kvypnuto je 5,6 a 0,038 s−1 a K.Ó2 (i.E. O.2-afinita vyplývající z kna/kvypnuto) je 23 a 1800 mM−1, resp.[11] To naznačuje, že sójový Lb může fungovat jako O2- skladování nebo - transportní protein a že funkce rýže Phytogb1 (a dalších hexa-souřadnicových fytogbs) může být jiná než O2- transport, protože vysoká afinita tohoto proteinu k O2 výsledky z extrémně nízké kvypnuto konstantní.[12]
Syntéza a postulované funkce
Stejně jako ostatní globiny, penta-souřadnice Phytogbs reverzibilně váží a transportují O2. Funkci Lbs v uzlinách objasnili v roce 1974 Wittenberg, Appleby a další.[13] V uzlinách je koncentrace Lbs velmi vysoká, protože odpovídají ~ 30% celkových rozpustných proteinů. Zdánlivá funkce Lbs v uzlících je usnadnit difúzi O2 na dýchající bakteroidy pro fixace dusíkem. Zároveň Lb přispívá k udržení nízké O2-úrovně (~ 10 nM), aby se zabránilo deaktivaci O2-citlivý dusičnan který fixuje atmosférický dusík.[14]
Fytogby dále vážou další plynné ligandy, zejména oxid dusnatý (NO), a vykazují NO dioxygenáza aktivita.[15] Práce Hilla a spolupracovníků během posledních ~ 15 let ukázala, že hladiny endogenního NO se mění s koncentrací Phytogbs1 v transgenní kukuřici a vojtěšce.[16] Na základě těchto pozorování tito autoři navrhli, že funkcí okysličených Phytogbs je modulovat hladiny NO prostřednictvím aktivity NO dioxygenázy a nepřímo regulovat širokou škálu buněčných funkcí, které jsou modulovány hladinami NO. Okysličené fytoglobiny třídy 1 reagující s NO za vzniku dusičnanů představují hlavní mechanismus, kterým je NO v rostlinách vychytáván. Cyklus zahrnující nitrátreduktázu, redukci dusitanu na NO, vychytávání NO fytoglobinem byl definován jako cyklus fytoglobin-NO[17]. Jeho provoz vede k udržení redoxního a energetického stavu během hypoxie a vede ke snížené produkci ethanolu a kyseliny mléčné.[18]
Phytogbs0, 1, 2 a 3 jsou syntetizovány ve velmi nízkých koncentracích v různých (embryonálních a vegetativních) rostlinných orgánech.[19][20][21] Koncentrace fytogbs se však zvyšuje u rostlin vystavených specifickým stresovým podmínkám, jako jsou záplavy[22] a omezení světla.[23] Některé fytogby byly proto považovány za proteiny reagující na stres rostlin.
Vývoj
Části tohoto článku (ty, které se týkají fylogeneze Becana odmítá společný původ pro Lbs a SymPhytogbs ve Phytogb1) je třeba aktualizováno. (Listopad 2020) |
Práce Arredondo-Petera, Vinogradova a spolupracovníků objasnila hlavní události, ke kterým mohlo dojít během vývoje Phytogbs.[24][25][26] Ačkoli se Phytogbs vyvinuly ze společného předka předcházejícího vzniku eukaryoty (i.E. v eubakterie v rostlinách byly pro Phytogbs identifikovány dvě evoluční linie: linie (3 / 3-) Phytogb0, 1 a 2 a SymPhytogb a Lb a linie (2 / 2-) Phytogb3. Zdá se, že Phytogbs0 byli předchůdci Phytogbs1 a 2 a SymPhytogbs a Lbs se vyvinuli z předchůdce Phytogb1 jako adaptace na symbiotickou fixaci dusíku. Srovnávací analýza mechu Phytogb0 s rýží Phytogb1 a strukturou sóji Lb odhalila, že hlavní evoluční změny, které pravděpodobně nastaly během vývoje Phytogbs0, 1 a 2, SymPhytogbs a Lbs byly (i) přechod hexa-souřadnice na penta-souřadnice v hemu skupina, (ii) zmenšení délky na CD-smyčce a N- a C-koncích a (iii) zhutnění proteinu do globulární struktury. Struktura globinové domény z Phytogbs3 je navíc velmi podobná struktuře bakteriálních zkrácených globinů, což naznačuje, že 2/2-skládání se zachovalo během vývoje Phytogbs3. Avšak globinová doména z rostlin Phytogbs3 je lemována sekvencemi extraaminokyselin, které pravděpodobně vznikly v předchůdci rostlin Phytogbs3. Funkční a evoluční význam sekvencí extra-aminokyselin ve struktuře Phytogbs3 stále není znám.
Technologické aplikace
Fytogby jsou potenciálně užitečné pro řadu biotechnologických aplikací. Například krátce po objevu rýže Phytogbs1 bylo zamýšlenou aplikací použití těchto proteinů jako Ó2-senzory v elektronických zařízeních. Rovněž byla navržena nadměrná exprese fytogbs jako strategie ke zvýšení tolerance plodin ke konkrétním stresovým podmínkám, jako je například záplava plodin [27] [28], protože spojuje signál předčasného zaplavení ethylen do málo kyslíku reakce rostlin [29][30]. Nedávno byly Phytogbs považovány za kandidáty na vývoj krevní náhrady a jako přísady v vegetariánské hamburgery.
Poznámky
Reference
- ^ Kubo H., Uber hamoprotein aus den wurzelknollchen von leguminosen, Acta Phytochim. (Tokio), 11 (1939) 195-200.
- ^ Vainshtein B. K., Harutyunyan E. H., Kuranova I. P., Borisov V. V., N.I. Sosfenov, Pavlovsky A. G., Grebenko A. I., Konareva N. V., Structure of leghaemoglobin from lupine root nodules at 5 A resolution., Nature, 254 (1975) 163-164
- ^ A b C d E F Becana, Manuel; Yruela, Inmaculada; Sarath, Gautam; Catalán, Pilar; Hargrove, Mark S. (září 2020). „Rostlinné hemoglobiny: cesta od jednobuněčných zelených řas k cévnatým rostlinám“. Nový fytolog. 227 (6): 1618–1635. doi:10,1111 / nph.16444.
- ^ A b C d Hill R., Hargrove M. S., Arredondo-Peter R (2016). „Fytoglobin: nová nomenklatura pro rostlinné globiny přijatá komunitou globinu na XVIII. Konferenci o proteinech vázajících a snímajících kyslík v roce 2014“. F1000Výzkum. 5: 212. doi:10.12688 / F1000Research.8133.1.CS1 maint: používá parametr autoři (odkaz)
- ^ Martínez-Ocampo F., Vázquez-Limón C., Arredondo-Peter R., Detekce, amplifikace PCR a charakterizace 4 intron obsahujícího nesymbiotického genu hemoglobinu z mechu Physcomitrella patens., Global J. Biochem., 3 (2012) 12
- ^ Hardison R. C., Stručná historie hemoglobinů: rostlin, zvířat, protistů a bakterií., Proc. Natl. Acad. Sci. USA., 93 (1996) 5675-5679
- ^ Go M., Korelace DNA exonových oblastí s proteinovými strukturními jednotkami v hemoglobinu., Nature, 291 (1981) 90-92.
- ^ Hyldig-Nielsen J. J., Jensen E., Paludan K., Wiborg O., Garret R., Joergersen P. L., Marker K. A., The primary structures of two leghemoglobin gens from soybean, Nucleic Acids Res., 10 (1982) 689-701.
- ^ Hargrove M., Brucker E. A., Stec B., Sarath G., Arredondo-Peter R., Klucas R. V., Olson J. S., Philips J. G. N., Krystalová struktura nesymbiotického hemoglobinu., Structure, 8 (2000) 1005-1014.
- ^ Reeder B. J., Hough M. A., Struktura nonsymbiotického rostlinného hemoglobinu třídy 3 z Arabidopsis thaliana odhaluje nové N-koncové šroubovité prodloužení., Acta Crystallogr., D70 (2014) 1411-1418.
- ^ Arredondo-Peter R., Hargrove M. S., Moran J. F., Sarath G., Klucas R. V., Plant hemoglobins, Plant Physiol., 118 (1998) 1121-1126.
- ^ Arredondo-Peter R., Hargrove M. S., Sarath G., Moran J. F., Lohrman J., Olson J. S., Klucas R. V., Rice hemoglobins: genové klonování, analýza a kinetika vazby kyslíku rekombinantního proteinu syntetizovaného v Escherichia coli., Plant Physiol., 115 (1997) 1259-1266
- ^ Wittenberg J. B., Bergersen F. J., Appleby C. A., Turner G. L., Facilitated kyslíková difúze: role leghemoglobinu při fixaci dusíku bakteroidy izolovanými z uzlíků sójových kořenů., J. Biol. Chem., 249 (1974) 4057-4066
- ^ Appleby C. A., Leghemoglobin a Rhizobium dýchání., Annu. Rev. Plant Physiol., 35 (1984) 443-478
- ^ Smagghe B. J., Trent J. T., Hargrove M. S., NO aktivita dioxygenázy v hemoglobinu je všudypřítomná in vitro, ale omezeno redukcí in vivo., PLoS One, 3 (2008) doi: 10,1371 / journal.pone.0002039.
- ^ Hill R. D., Nesymbiotické hemoglobiny - Co se děje mimo zachycování oxidu dusnatého ?, AoB Plants, Pls004 (2012) doi: 10,1093 / aobpla / pls1004
- ^ Igamberdiev, A.U .; Baron, K .; Manac'h-Little, N .; Stoimenova, M .; Hill, RD (2005). „Cyklus hemoglobin / oxid dusnatý: účast na povodňových stresech a účinky na hormonální signalizaci“. Annals of Botany. 96 (4): 557–564. doi:10.1093 / aob / mci210. ISSN 0305-7364. PMC 4247025. PMID 16027133.
- ^ Igamberdiev, A.U .; Hill, RD (2004). „Dusičnany, NO a hemoglobin v adaptaci rostlin na hypoxii: alternativa ke klasickým způsobům fermentace“. Journal of Experimental Botany. 55 (408): 2473–2482. doi:10.1093 / jxb / erh272. ISSN 0022-0957. PMID 15448180.
- ^ Garrocho-Villegas V., Arredondo-Peter R., Molekulární klonování a charakterizace mechu (Ceratodon purpureus) nesymbiotický hemoglobin poskytuje pohled na časný vývoj rostlinných nesymbiotických hemoglobinu., Mol. Biol. Evol., 25 (2008) 1482-1487.
- ^ Ross EJH, Shearman L., Mathiesen M., Zhou J., Arredondo-Peter R., Sarath G., Klucas RV, nesymbiotické hemoglobiny jsou syntetizovány během klíčení a při diferenciaci typů buněk., Protoplasma, 218 (2001) 125 -133.
- ^ Watts R. A., Hunt P. W., Hvitved A. N., Hargrove M. S., Peacock W. J., Dennis E. S., Hemoglobin z rostlin homologních se zkrácenými hemoglobiny mikroorganismů., Proc. Natl. Acad. Sci. USA., 98 (2001) 10119-10124.
- ^ Taylor E. R., Nie X. Z., MacGregor A. W., Hill R. D., Cereální hemoglobinový gen je exprimován v semenných a kořenových tkáních za anaerobních podmínek, Plant Mol. Biol., 24 (1994) 853-862
- ^ Lira-Ruan V., Sarath G., Klucas R. V., Arredondo-Peter R., Syntéza hemoglobinů v rýži (Oryza sativa var. Jackson) rostliny rostoucí za normálních a stresových podmínek., Plant Sci., 161 (2001) 279-287.
- ^ Vázquez-Limón C., Hoogewijs D., Vinogradov S. N., Arredondo-Peter R., Evoluce hemoglobinů suchozemských rostlin, Plant Sci., 191-192 (2012) 71-81.
- ^ Vinogradov S. N., Fernández I., Hoogewijs D., Arredondo-Peter R., Fylogenetické vztahy hemoglobinů 3/3 a 2/2 v genomech Archaeplastida k bakteriálním a jiným eukaryotickým hemoglobinům, Mol. Plant, 4 (2011) 42-58.
- ^ Vinogradov S. N., Hoogewijs D., Arredondo-Peter R., What are the origin and phylogeny of plant hemoglobins ?, Commun. Integrat. Biol., 4 (2011) 443-445.
- ^ Andrzejczak, OA; Havelund, JF; Wang, WQ; Kovalchuk, S; Hagensen, CE; Hasler-Sheetal, H; Jensen, ON; Rogowska-Wrzesinska, A; Møller, IM; Hebelstrup, KH (24. února 2020). „Hypoxický protein a metabolom z ječmene (Hordeum vulgare L.) sa bez použití fytoglobinu ". International Journal of Molecular Sciences. 21 (4): 1546. doi:10,3390 / ijms21041546. PMID 32102473.
- ^ Mira, Mohamed M .; Hill, Robert D .; Stasolla, Claudio (listopad 2016). „Fytoglobiny zlepšují hypoxický růst kořenů zmírňováním apikální meristemové buněčné smrti“. Fyziologie rostlin. 172 (3): 2044–2056. doi:10.1104 / str. 16.01150. PMC 5100795. PMID 27702845.
- ^ Hartman, Johannes Gerardus Wilhelmus (2020-01-22). Signál předčasného zaplavení ethylen aklimatizuje rostliny, aby přežily stres s nízkým obsahem kyslíku. Universiteit Utrecht. hdl:1874/387763. ISBN 978-90-393-7231-9.
- ^ Hartman, S; Liu, Z; van Veen, H; Vicente, J; Reinen, E; Martopawiro, S; Zhang, H; van Dongen, N; Bosman, F; Bassel, GW; Visser, EJW; Bailey-Serres, J; Theodoulou, FL; Hebelstrup, KH; Gibbs, DJ; Holdsworth, MJ; Sasidharan, R; Voesenek, LACJ (5. září 2019). „Vyčerpání oxidu dusnatého zprostředkované ethylenem před adaptuje rostliny na stres hypoxií“. Příroda komunikace. 10 (1): 4020. Bibcode:2019NatCo..10.4020H. doi:10.1038 / s41467-019-12045-4. PMC 6728379. PMID 31488841.