Dyrosauridae - Dyrosauridae
| Dyrosauridae | |
|---|---|
 | |
| Lebka dyrosaurida Arambourgisuchus khouribgaensis | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Třída: | Reptilia |
| Podřád: | †Tethysuchia |
| Rodina: | †Dyrosauridae de Stefano, 1903 |
| Rody | |
Dyrosauridae je rodina zaniklý neosuchian krokodýli který žil z Pozdní křída (Maastrichtian ) do Eocen. Fosílie dyrosauridů jsou celosvětově distribuovány a byly nalezeny v Africe, Asii, Evropě, Severní Americe a Jižní Americe. V současné době je známo více než tucet druhů, které se velmi liší celkovou velikostí a lebečním tvarem. Všichni byli pravděpodobně vodní, s druhy obývajícími jak sladkovodní, tak mořské prostředí. Dyrosauridové žijící v oceánu byli mezi několika mořskými plazy, kteří přežili Událost vyhynutí křída – paleogen.
Dyrosauridi byli skupina převážně mořských čtyřlístků podobných krokodýlům s dlouhými čelistmi, dlouhých až 6 metrů (20 stop).[1] Na základě důkazů o kostní tkáni se předpokládá, že rostou pomalu[2] mořští živočichové na pobřeží s uzavřenými čelistmi,[2] umí plavat i chodit na souši. Vnější nosní dírky na zadním konci čenichu a vnitřní naris v jeho pterygoidu naznačovaly zvyk lovit při plavání s temenem hlavy nad vodou, což mu umožňovalo dýchat při pronásledování kořisti.[1]
Přehled

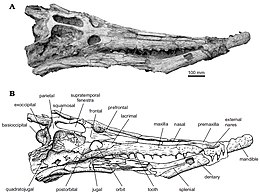
Dyrosaurids je známo, že mají velmi charakteristický tvar lebky s dlouhým a tenkým čenichem, který je přibližně 68% z celkové délky lebky. Nejpřednější část lebky a čenichu je externí naris následovaný dozadu dvěma premaxillae kosti, dokud nedosáhnou dvou maxillae kosti oddělené jedinou nosní kost.[1]
Typický pro dyrosauridy je jediný nosní prvek s charakteristickou sbírkou malých jamek a konstantní šířkou, dokud se nerozšíří, aby se dotkl slzný kosti, poté se zužuje na krátkou vzdálenost, dokud nedosáhne hranice frontálních a prefrontálních.[1]

Dyrosaury mají premaxilla s mělkými jámami, které sahají dozadu ke třetím čelistním alveolům. Existují dva premaxillae, které jsou ve srovnání s maxilárními kostmi úzké a táhnou se ve dvou dlouhých horní čelist kosti oddělené jedinou nosní kostí. Poslední premaxilla a první maxilla jsou široce odděleny čtvrtým zubem zubu. Alveoly jsou vpředu široce rozmístěny a prostor mezi nimi se zmenšuje dozadu z patnáctého alveolu s konstantním průměrem.[1]
Maxilla je dlouhá (přibližně dvaapůlnásobek délky jugal ) a tvoří většinu bočního okraje lebky.[3] Podle Jouveho a Barbosy a možná v závislosti na věku zvířete má každá maxilla 13-19 zubů.[3]
Důležitá vlastnost dyrosauridu chrup jsou hluboké okluzní jámy, přítomné zejména v zadní oblasti čelistí, které se vpředu méně zvýrazňují. Jámy svědčí o vzájemně propojené uzavřené čelisti pro dyrosauridy, protože jámy umožňují střídání horní a dolní čelisti. I když jsou stále přítomny v dyrosaurus phosphaticus, tato vlastnost hlubokých okluzních jám se stává méně rozvinutou a širokou.[3] Zuby dyrosaura jsou homodontní, kuželovité, dlouhé a štíhlé, s asymetricky labiálními a lingválními povrchy. Zadní zuby jsou kratší a stlačenější než přední zub, což naznačuje, že velikost zubu se zmenšila před zadním.

Dyrosaury mají čenich k délce lebky asi 68% a rod Rhabdognathus a Atlantosuchus, Dyrosaurus a Arambourgisuchus mají největší podíl čenichu ze všech dyrosauridů. Délka čenichu se dříve používala k navázání dyrosauridových vztahů, přičemž prodloužení čenichu považovala za „rozvinutější“ charakter. To nebylo v souladu s Jouveho závěrem, že nejdelší čenich je ve skutečnosti primitivní podmínkou, takže kratší nebo delší čenich se v dyrosauridové evoluci objeví samostatně alespoň čtyřikrát.[4]
Paleobiogeografie
Dyrosauridi byli kdysi považováni za africkou skupinu, ale objevy od 2000u ukazují, že obývali většinu kontinentů.[5] Ve skutečnosti bazální formy naznačují, že jejich kolébkou mohla být Severní Amerika.
Všeobecné
| Rod | Postavení | Stáří | Umístění | Popis | snímky |
|---|---|---|---|---|---|
| Acherontisuchus | Platný | Paleocen | Velký, dlouho čichající sladkovodní dyrosaurid z Cerrejónské souvrství | ||
| Aigialosuchus | Platný | Campanian | Mořský dyrosaurid z Kristianstad Basin | ||
| Anthracosuchus | Platný | Paleocen | Sladkovodní dyrosaurid s krátkým čenichem z Cerrejónské souvrství | ||
| Arambourgisuchus | Platný | Paleocen | Dlouho čenichový mořský dyrosaurid | ||
| Atlantosuchus | Platný | Paleocen | Mořský dyrosaurid s dlouhým čenichem s nejdelší délkou čenichu v poměru k velikosti těla jakéhokoli dyrosauridu | ||
| Cerrejonisuchus | Platný | Paleocen | Malý, krátkosrstý sladkovodní dyrosaurid z Cerrejónské souvrství | ||
| Chenanisuchus | Platný | Maastrichtian -Paleocen | Rod pokrývá K-Pg hranice |  | |
| Congosaurus | Platný | Paleocen | |||
| Dyrosaurus | Platný | Eocen | Velký mořský dyrosaurid s dlouhým nosem |  | |
| Fortignathus | Platný | Albian –Cenomanský | Dyrosaurid z Echkarské souvrství. | ||
| Guarinisuchus | Platný | Paleocen | Mořský dyrosaurid |  | |
| Hyposaurus | Platný | Maastrichtian -Paleocen | Bylo pojmenováno pět druhů, nejvíce ze všech rodů dyrosauridů; rod pokrývá K-Pg hranice | ||
| Phosphatosaurus | Platný | Eocen | Velký mořský dyrosaurid s dlouhým čenichem s tupými zuby a špičkou čenichu ve tvaru lžíce | ||
| Rhabdognathus | Platný | Maastrichtian -Paleogen | Velký mořský dyrosaurid s dlouhým nosem; rod pokrývá K-Pg hranice | ||
| Sabinosuchus | Platný | Maastrichtian | Dyrosaurid z Escondido formace. | ||
| Sokotosaurus | Junior synonymum | Junior synonymum pro Hyposaurus | |||
| Sokotosuchus | Platný | Maastrichtian | Dlouho čenichový mořský dyrosaurid | ||
| Tilemsisuchus | Platný | Eocen |
Fylogeneze
Jouve et al. (2005) diagnostikují Dyrosauridae jako a clade na základě následujících sedmi synapomorfie nebo sdílené postavy:
- Posteromediální křídlo retroartikulárního výběžku dorzálně situované ventrálně na retroartikulárním výběžku
- Týlní tuberosity malé
- Exoccipitální se z velké části podílí na týlní kondyl
- Supratemporální fenestra předozadně silně protáhlá
- Symphysis asi tak široký jako vysoký
- Quadratojugal se z velké části podílí na lebečním kondylu pro artikulaci s čelistí
- 4 premaxilární zuby
Níže je a kladogram po Jouve et al. (2005) ukazující fylogenetické vztahy Dyrosauridae a dalších blízce příbuzných neosuchians:
| Neosuchia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Kompozitní kladogram pro Dyrosauridae (od Jouve et al. 2008 a Barbosa et al. 2008):
| Dyrosauridae |
Dyrosauridae incertae sedis:Tilemsisuchus | |||||||||||||||||||||||||||||||||||||||||||||||||||
Cladogram po Hastingsovi et al. (2011) ukazující geografické výskyty taxonů:[6]
| Neosuchia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Analýza naznačuje, že nejbližší příbuzní dyrosauridů jsou Sarkosuchus a Terminonaris.
Paleobiologie
Růst

Důkazy o semi-vodním životě dyrosauridů pocházejí z pečlivé analýzy struktury kostí. U vodních tetrapodů mohou nastat dva typy strukturní organizace kostí: osteoporotický nebo pachyostotický. Osteoporotická kost je houbovitá a porézní, zatímco pachyostotická zahrnuje zvýšení kostní hmoty. Hubovitá / porézní kost, jako je osteoporotika, je spojena s rychlejším plaváním a lepší manévrovatelností ve vodě kvůli snížení kostní tkáně, mnoho existujících kytovců a mořských želv má osteoporotickou kost, což jim umožňuje být dobrými plavci. Pachyostotická kost je obecný / lokální nárůst kosterní hmoty, který může být způsoben osteosklerózou (vnitřní zhutnění kosti), pachyostózou (hyperplazie kompaktních kortik) nebo pachyeosklerózou (kombinace těchto dvou). Výzkum kosti dyrosaura provedený Rafaelem Césarem Limou Pedroso de Andrade Julianou Manso Sayao odhalil, že tato rodina měla osteoporotickou kostní tkáň svědčící o rychlém plavání ekologie stejně jako některé osteoskleróza což je součást pachyostotické kostní tkáně. Osteoporóza je spojena s plně vodním životním stylem, zatímco pachyostotika není zcela vodní, ale je spojena s ekologií rychlého plavání. Proto jsou dyrosauri polovodními rychlými plavci, což naznačuje jejich struktura kostí. Dalším důkazem pro blízký břeh je polo-vodní životní styl, kde se nacházejí fosilie, často v přechodných mořských sedimentech.[2] -použití plavání v osové frekvenci (které používají především existující krokodýli) s větším vlnivý pohyb a frekvence ocasu díky vysoce vyvinutému svalstvo umožňující silnější dopředný tah.[2]
Dyrosauridy mají tkáňový vzor, který svědčí o zvířeti s pomalým růstem, které bylo určeno pečlivou analýzou pravého stehenní kost a vlevo holenní kost. V levé holenní kosti měla kůra lamelární zonální kost s pěti liniemi zastaveného růstu (LAG), které byly rozmístěny 300 mm od sebe, a také jasné vaskulární sítě primárních osteonů, které snižovaly hustotu směrem k membráně (periostiálne). Pravá femur měla dvojité LAG a EFS později, stejně jako sekundární osteony vyskytující se v hluboké kůře a spongiose. Tento vzor růstu tkáně je běžnou charakteristikou pomalu rostoucích zvířat.[2]
Místo výskytu
Dyrosauridy se nacházejí v přechodných mořských sedimentech od pozdní křídy po nižší eocén.[2] Tato rodina je známá hlavně z ložisek Maastrichthian v New Jersey a od pozdní křídy po ranou paleogenní horninu z moře Tethys v severní a západní Africe.[2] Fosílie byly také nalezeny z paleocénu a eocénu v Pákistánu, stejně jako v Jižní Americe, Brazílii, Indii, jižní Asii a na pobřeží. Obecně se dyrosaurids získávají z pobřežních ložisek a ložisek v ústí řek v severní Africe a na Středním východě, což potvrzuje jejich existenci jako polovodních živočichů.[7]
Dyrosauridae měli během raného období největší taxonomickou rozmanitost Paleogen ale zdá se, jako by clade dokázal během pozdní křídy získat větší a rozšířenější geografické rozložení. Nejstarší záznamy o dyrosauridech jsou buď v Africe, nebo v její blízkosti, s fragmentárními výskyty z Cenomanský Súdánu a Portugalska a několika dalšíchMaastrichtian „Pozdně křídové objevy v Egyptě. Později Maastrichtianem ze Severní Ameriky se záznam dyrosauridů stal úplnějším vytvořením rozšířené distribuce, která se zdá být udržována prostřednictvím paleocenu a eocénu.[8]
Dyrosaurids byly také nalezeny z non-mořských sedimentů. Na severu Súdán, dyrosauridy jsou známy z říční ložiska, což naznačuje, že žili v říčním prostředí.[9] Kosti z neurčitých dyrosauridů byly nalezeny ve vnitrozemských ložiscích v Pákistán také. Některé dyrosauridy, například ty z Umm Himar formace v Saudská arábie, obydlený ústí prostředí poblíž pobřeží. Nedávno pojmenovaní dyrosaurids Cerrejonisuchus a Acherontisuchus byly získány z Cerrejónské souvrství na severozápadě Kolumbie, o kterém se předpokládá, že představuje přechodné mořské a sladkovodní prostředí obklopené deštnými pralesy více do vnitrozemí než do ústí řek formace Umm Himar.[10] Cerrejonisuchus a Acherontisuchus žil v neotropickém prostředí v době, kdy byly globální teploty mnohem teplejší než jsou dnes.[11][12]
Reprodukce
V roce 1978 bylo navrženo, aby dyrosauridové žili jako dospělí v oceánu, ale rozmnožovali se ve vnitrozemských sladkovodních prostředích. Pozůstatky drobných dyrosauridů z Pákistánu byly interpretovány jako mladiství. Jejich přítomnost ve vnitrozemských ložiscích byla považována za důkaz, že se dyrosaurids vylíhla daleko od oceánu.[13] Z dolní eocénské nádrže Oulad Abdoun je velmi málo juvenilních dyrosauridů, ale mnoho podobných dospělých exemplářů. To podpořilo předpoklad, že mladiství žili ve sladkovodním prostředí a dospělí žili v mořském prostředí.[14] V poslední době však velcí a plně zralí dyrosauridi formace Cerrejón ukázali, že někteří dyrosauridové žili celý život ve vnitrozemských prostředích a nikdy se nevrátili na pobřeží.[6]
Reference
- ^ A b C d E Jouve, Stephanie. Nový popis lebky Dyrosaurus phosphaticus (Thomas, 1893) (Mesoeucrocodylia: Dyrosauridae) ze spodního eocénu severní Afriky. 2005. Citováno 2. března 2015 z http://cjes.nrc.ca Archivováno 04.06.2002 na Wayback Machine.
- ^ A b C d E F G Andrade RCLPd, Saya ̃o JM (2014) Paleohistologie a důsledky životního stylu dyrosaurida (Archosauria: Crocodylomorpha) z pánve Paraíba (severovýchodní Brazílie). PLOS ONE 9 (7): e102189. doi: 10,1371 / journal.pone.0102189
- ^ A b C Antonio Barbosa, José; Wilhelm Armin Kellner, Alexander. Somália Sales Viana, Maria. Nový krokosylomid dyrosauridů a důkazy faunálního obratu při přechodu K – P v Brazílii. 2008. Sborník Královské společnosti. Staženo z http://rspb.royalsocietypublishing.org/
- ^ Jouve, Stephanie; Bouya, Baadi; Amaghzaz, Mbarek. DLOUHODOBÝ DYROSAURID (CROCODYLIFORMES, MESOEUCROCODYLIA) Z MAROKOU PALEOCENY: FYLOGENETICKÉ A PALAEOBIOGEOGRAFICKÉ DOPADY. 2008. Paleontologie, roč. 51, část 2. str. 281-294.
- ^ Jouve et al. (2008)
- ^ A b Hastings, A.K., Bloch, J. a Jaramillo, C.A. (2011). „Nový longirostrinový dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) z paleocenu v severovýchodní Kolumbii: biogeografické a behaviorální důsledky pro nový svět dyrosauridae“ (PDF). Paleontologie. 54 (5): 1095–116. doi:10.1111 / j.1475-4983.2011.01092.x. Archivovány od originál (PDF) dne 31. března 2012. Citováno 14 zář 2011.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Matthew C. Lamanna, Joshua B. Smith, Yousry S. Attia a Peter Dodson Zdroj: Journal of Vertebrate Paleontology, Vol. 24, č. 3 (10. září 2004), str. 764-768 Publikováno: Taylor & Francis, Ltd. jménem Společnosti paleontologie společnosti obratlovců Stabilní URL: https://www.jstor.org/stable/4524765 Přístup: 04-03-2015 17:11 UTC.
- ^ Ashu Khosla, Joseph J. W. Sertich, Guntupalli V. R. Prasad a Omkar Verma Zdroj: Journal of Vertebrate Paleontology, Vol. 29, č. 4 (12. prosince 2009), str. 1321–1326 Publikováno: Taylor & Francis, Ltd. jménem Společnosti paleontologie společnosti obratlovců Stabilní URL: https://www.jstor.org/stable/20627142. Přístup: 03-03-2015 07:01 UTC
- ^ Buffetaut, E .; Bussert, R .; Brinkmann, W. (1990). „Nová nemořská obratlovcová fauna ve svrchní křídě severního Súdánu“. Berliner Geowissenschaftlische Abhandlungen. 120: 183–202.
- ^ Hastings, A. K; Bloch, J. I .; Cadena, E. A .; Jaramillo, C. A. (2010). „Nový malý krátkosrstý dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) z paleocénu v severovýchodní Kolumbii“. Časopis paleontologie obratlovců. 30 (1): 139–162. doi:10.1080/02724630903409204. S2CID 84705605.
- ^ Head, J. J .; Bloch, J. I .; Hastings, A. K .; Borque, J. R .; Cadena, E. A .; Herrera, F. A .; Polly, P. D .; Jaramillo, C. A. (2009). „Obří boidský had z paleocénové neotropiky odhaluje teplejší minulé rovníkové teploty“. Příroda. 457 (7230): 715–717. Bibcode:2009 Natur.457..715H. doi:10.1038 / nature07671. PMID 19194448. S2CID 4381423.
- ^ Kanapaux, B. (2. února 2010). „Výzkumníci UF: Starověký krokodýl relativní pravděpodobný zdroj potravy pro Titanoboa“. University of Florida News. Archivovány od originál 8. června 2010. Citováno 3. února 2010.
- ^ Buffetaut, E. (1978). "Krokodýlí pozůstatky z eocénu Pákistánu". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 156: 262–283.
- ^ Jouve, Stephanie; Bardet, Nathalie; Jalil, Nour-Eddine; Superbiola, Xabier Pereda; Bouya, Baadi a Amagzaz, Mbarek. 2008. Nejstarší africký krokodýl: fylogeneze, paleobiogeografie a rozdílné přežití mořských plazů přes hranici křídové a terciární. Časopis paleontologie obratlovců 28 (2): 409–421, červen 2008
- Barbosa, J.A., Kellner, A.W.A. a Viana, M.S.S. (2008). Nový krokosylomid dyrosauridů a důkazy faunálního obratu při přechodu K – P v Brazílii. Sborník Královské společnosti B: Biologické vědy: Firstcite
- Buffetaut, E. (1985). L'evolution des crocodiliens. Les animaux disparus-Pour la science, Paříž 109.
- Jouve, S .; Bouya, B .; Amaghzaz, M. (2008). „Dlouho čenichový dyrosaurid (Crocodyliformes, Mesoeucrocodylia) z paleocénu v Maroku: fylogenetické a paleogeografické důsledky“. Paleontologie. 51 (2): 281–294. doi:10.1111 / j.1475-4983.2007.00747.x.
- Jouve, S .; Iarochène, M .; Bouya, B .; Amaghzaz, M. (2005). „Nový krokodýlí dyrosaurid z marockého paleocénu a fylogenetická analýza Dyrosauridae“. Acta Palaeontologica Polonica. 50 (3): 581–594.
externí odkazy
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||

