Solitonový model v neurovědě - Soliton model in neuroscience
Tento článek má několik problémů. Prosím pomozte vylepši to nebo diskutovat o těchto otázkách na internetu diskusní stránka. (Zjistěte, jak a kdy tyto zprávy ze šablony odebrat) (Zjistěte, jak a kdy odstranit tuto zprávu šablony)
|
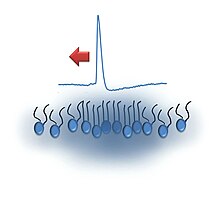
The solitonová hypotéza v neurovědy je Modelka který tvrdí, že vysvětluje jak akční potenciály jsou zahájeny a prováděny společně axony na základě termodynamické teorie šíření nervových pulsů.[1] Navrhuje, aby se signály šířily podél buňky membrána ve formě určitých druhů osamělý zvuk (nebo hustota ) pulsy, které lze modelovat jako solitony. Tento model je navržen jako alternativa k Hodgkin – Huxleyův model[2] ve kterém akční potenciály: napěťově řízené iontové kanály v membráně otevřete a nechejte sodík ionty pro vstup do buňky (vnitřní proud). Výsledný pokles membránového potenciálu otevírá blízké napěťově řízené sodíkové kanály, čímž se šíří akční potenciál. Transmembránový potenciál je obnoven opožděným otevřením draslíkových kanálů. Zastánci hypotézy Soliton tvrdí, že energie se během šíření hlavně zachovává, kromě ztrát rozptylem; Změřené změny teploty jsou zcela nekonzistentní s Hodgkin-Huxleyovým modelem.[3][4]
Solitonový model (a zvuk vlny obecně) závisí na adiabatickém šíření, při kterém je energie poskytovaná u zdroje excitace přenášena adiabaticky médiem, tj. plazmatickou membránou. Měření teplotního impulsu a údajné nepřítomnosti uvolňování tepla během akčního potenciálu[5][6] byly základem návrhu, že nervové impulsy jsou adiabatickým jevem podobně jako zvukové vlny. Synapticky vyvolané akční potenciály v elektrickém orgánu elektrického úhoře jsou spojeny s podstatnou pozitivní (jedinou) produkcí tepla následovanou aktivním ochlazením na teplotu okolí.[7] V čichovém nervu garfish je akční potenciál spojen s dvoufázovou změnou teploty; Existuje však čistá výroba tepla.[8] Tyto publikované výsledky jsou nekonzistentní s Hodgkin-Huxleyovým modelem a autoři interpretují svou práci ve smyslu tohoto modelu: Počáteční sodíkový proud uvolňuje teplo při vybití kapacity membrány; teplo se absorbuje během dobíjení kapacity membrány, když se ionty draslíku pohybují s jejich koncentračním gradientem, ale proti potenciálu membrány. Tento mechanismus se nazývá „teorie kondenzátoru“. Další teplo může být generováno změnami konfigurace membrány způsobenými změnami membránového potenciálu. Zvýšení entropie během depolarizace by uvolnilo teplo; zvýšení entropie během repolarizace by absorbovalo teplo. Jakékoli takové entropické příspěvky jsou však nekompatibilní s Hodgkinovým a Huxleyovým modelem[9]
Dějiny
Ichiji Tasaki propagoval termodynamický přístup k fenoménu šíření nervových pulsů, který identifikoval několik jevů, které nebyly zahrnuty do Hodgkin – Huxleyův model.[10] Spolu s měřením různých neelektrických složek nervového impulsu zkoumal Tasaki fyzikální chemii fázové přechody v nervových vláknech a jeho význam pro šíření nervových pulsů. Na základě Tasakiho díla navrhl Konrad Kaufman zvukové vlny jako fyzický základ pro šíření nervových pulsů v nepublikovaném rukopisu.[11] Základní myšlenkou jádra solitonového modelu je vyvážení vnitřní disperze dvourozměrných zvukových vln v membráně nelineárními elastickými vlastnostmi poblíž fázového přechodu. Počáteční impuls může za takových okolností získat stabilní tvar, obecně známý jako osamělá vlna.[12] Solitony jsou nejjednodušším řešením souboru nelineárních vlnových rovnic, které tento jev řídí, a byly aplikovány na modelování nervových impulsů v roce 2005 Thomasem Heimburgem a Andrewem D. Jacksonem,[13][14][15] oba u Institut Nielse Bohra z Kodaňská univerzita. Heimburg stojí v čele institutu Membrane Biophysics Group. Skupina biologické fyziky Matthiase Schneidera studovala šíření dvourozměrných zvukových vln v lipidových rozhraních a jejich možnou roli v biologické signalizaci [16][17][18][19]
Odůvodnění
Model začíná pozorováním, že buněčné membrány mají vždy a bod mrazu (teplota, pod níž se konzistence mění z tekuté na gelovou), jen mírně pod tělesnou teplotou organismu, což umožňuje šíření solitonů. Akční potenciál putující podél smíšeného nervu má za následek mírné zvýšení teploty následované poklesem teploty.[20] Zastánci modelu Soliton tvrdí, že během celkového pulzu se neuvolňuje žádné čisté teplo a že pozorované změny teploty nejsou v souladu s Hodgkin-Huxleyovým modelem. To je však nepravdivé: model Hodgkin Huxley předpovídá dvoufázové uvolňování a absorpci tepla.[9] Kromě toho akční potenciál způsobuje mírné lokální zesílení membrány a sílu působící směrem ven;[21] tento efekt není předpovězen Hodgkin-Huxleyovým modelem, ale ani mu není v rozporu.
Solitonový model se pokouší vysvětlit elektrické proudy spojené s akčním potenciálem následovně: cestující soliton lokálně mění hustotu a tloušťku membrány, a protože membrána obsahuje mnoho nabitých a polární látek, bude to mít podobný elektrický účinek piezoelektřina. Ve skutečnosti se nyní ukázalo, že takové nelineární zvukové vlny existují na lipidových rozhraních, která vykazují povrchní podobnost s akčními potenciály (elektroopto-mechanická vazba, rychlosti, dvoufázový tvar pulzu, práh pro excitaci atd.).[17] Vlny dále zůstávají lokalizovány v membráně a nešíří se v okolí kvůli nesouladu impedance.[22]
Formalismus
Soliton představující akční potenciál nervů je řešením parciální diferenciální rovnice
![{frac {partial ^{2}Delta
ho }{partial t^{2}}}={frac {partial }{partial x}}left[left(c_{0}^{2}+pDelta
ho +qDelta
ho ^{2}
ight){frac {partial Delta
ho }{partial x}}
ight]-h{frac {partial ^{4}Delta
ho }{partial x^{4}}},](https://wikimedia.org/api/rest_v1/media/math/render/svg/49468442896ff1edd5f1297b1a2253d2dee7a782)
kde t je čas a X je poloha podél nervového axonu. Δρ je změna hustoty membrány pod vlivem akčního potenciálu, C0 je rychlost zvuku nervové membrány, p a q popsat povahu fázového přechodu a tím nelinearitu elastických konstant nervové membrány. Parametry C0, p a q jsou diktovány termodynamickými vlastnostmi nervové membrány a nelze je volně nastavovat. Musí být stanoveny experimentálně. Parametr h popisuje frekvenční závislost rychlosti zvuku membrány (disperzní vztah ). Výše uvedená rovnice neobsahuje žádné parametry přizpůsobení. Formálně souvisí s Boussinesqova aproximace pro solitony ve vodních kanálech. Řešení výše uvedené rovnice mají omezující maximální amplitudu a minimální rychlost šíření, která je podobná rychlosti pulzu v myelinovaných nervech. Podle omezujících předpokladů existují periodická řešení, která zobrazují hyperpolarizaci a refrakterní období.[23]
Úloha iontových kanálů
Zastánci solitonového modelu tvrdí, že vysvětluje několik aspektů akčního potenciálu, které nejsou vysvětleny Hodgkin-Huxleyovým modelem. Jelikož má termodynamickou povahu, neřeší vlastnosti jednotlivých makromolekul iontový kanál proteiny v molekulárním měřítku. Spíše se předpokládá, že jejich vlastnosti jsou implicitně obsaženy v makroskopických termodynamických vlastnostech nervových membrán. Solitonový model předpovídá fluktuace membránového proudu během akčního potenciálu. Tyto proudy mají podobný vzhled jako proudy uváděné pro proteiny iontových kanálů.[24] Předpokládá se, že jsou způsobeny póry lipidové membrány spontánně generovanými tepelnými výkyvy. Takové tepelné fluktuace vysvětlují specifickou iontovou selektivitu nebo specifický časový průběh reakce na změny napětí na základě jejich vlivu na makroskopické susceptibility systému.
Aplikace do anestézie
Autoři tvrdí, že jejich model vysvětluje dříve nejasný způsob působení mnoha anestetika. The Pozorování Meyer – Overton tvrdí, že síla široké škály chemicky rozmanitých anestetik je úměrná jejich síle lipid rozpustnost, což naznačuje, že nejednají vazbou na konkrétní bílkoviny jako jsou iontové kanály, ale místo toho rozpuštěním a změnou vlastností lipidové membrány. Rozpouštění látek v membráně snižuje bod tuhnutí membrány a výsledný větší rozdíl mezi tělesnou teplotou a bodem mrazu brání šíření solitonů.[25] Zvyšováním tlaku, snižováním pH nebo snížení teploty může být tento rozdíl obnoven zpět do normálu, což by mělo zrušit působení anestetik: to je skutečně pozorováno. Míra tlaku potřebného ke zrušení působení anestetika dané rozpustnosti v tucích lze vypočítat z modelu soliton a přiměřeně dobře souhlasí s experimentálními pozorováními.
Rozdíly mezi predikcemi modelu a experimentálními pozorováními
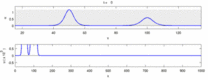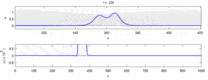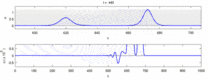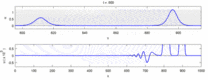
Následuje seznam některých sporů mezi experimentálními pozorováními a „solitonovým modelem“:
- Antidromická invaze soma z axonu
- Akční potenciál zahájený kdekoli na axonu bude cestovat antidromickým (zpětným) směrem k neuronové somě (tělu buňky) bez ztráty amplitudy a vytvoří akční potenciál s plnou amplitudou v somě. Vzhledem k tomu, že membránová plocha soma je řádově větší než plocha axonu, vyžaduje zachování energie snížení adiabatické mechanické vlny o amplitudu. Vzhledem k tomu, že absence výroby tepla je jedním z nárokovaných odůvodnění „solitonového modelu“, je v rámci tohoto modelu obzvláště obtížné to vysvětlit.[26][Citace je zapotřebí ]
- Perzistence akčního potenciálu v širokém teplotním rozsahu
- Důležitým předpokladem solitonového modelu je přítomnost fázového přechodu blízko teploty okolí axonu („formalismus“ výše). Potom by rychlá změna teploty mimo teplotu fázového přechodu nutně způsobila velké změny akčního potenciálu. Pod teplotou fázového přechodu by solitonová vlna nebyla možná. Přesto jsou akční potenciály přítomny při 0 ° C. Časový průběh je zpomalen způsobem předpovězeným měřenou kinetikou otevírání a zavírání iontových kanálů Hodgkin-Huxley.[27]
- Kolize
- Nervové impulsy cestující v opačných směrech se při srážce navzájem ničí.[28] Na druhou stranu mechanické vlny nevyhazují, ale procházejí navzájem. Navrhovatelé modelu Soliton se pokusili ukázat, že akční potenciály mohou projít kolizí;[29] kolizní zničení ortodromních a antidromní akční potenciály jsou v neurovědeckých laboratořích běžně pozorovaným jevem a jsou základem standardní techniky identifikace neuronů.[30] Solitony se navzájem protínají při kolizi (obrázek - „Kolize solitonů“), osamělé vlny se obecně mohou navzájem procházet, ničit nebo se odrážet[31] a solitony jsou jen zvláštním případem takových osamělých vln.[32]
- Iontové proudy pod napěťovou svorkou
- Napěťová svorka, kterou používali Hodgkin a Huxley (1952) (Hodgkin-Huxleyův model ) k experimentální pitvě akčního potenciálu v gigantickém axonu chobotnice, používá elektronickou zpětnou vazbu k měření proudu potřebného k udržení konstantního napětí membrány na předepsané hodnotě. Stříbrný drát, vložený do vnitřku axonu, nutí konstantní membránové napětí po celé délce axonu. Za těchto okolností neexistuje možnost cestování „soliton“. Jakékoli termodynamické změny se velmi liší od změn vyplývajících z akčního potenciálu. Měřené proudy přesto přesně reprodukují akční potenciál.[Citace je zapotřebí ]
- Jednokanálové proudy
- Technika svorky náplasti izoluje mikroskopickou náplast membrány na špičce skleněné pipety. Potom je možné zaznamenávat proudy z jednotlivých iontových kanálů. Neexistuje možnost šíření solitonů nebo termodynamických změn. Vlastnosti těchto kanálů (časová odezva na napěťové skoky, iontová selektivita) přesto přesně předpovídají vlastnosti makroskopických proudů měřených konvenční napěťovou svorkou.[33]
- Selektivní iontová vodivost
- Proud, který je základem depolarizace akčního potenciálu, je pro sodík selektivní. Repolarizace závisí na selektivním proudu draslíku. Tyto proudy mají velmi specifické reakce na změny napětí, které kvantitativně vysvětlují akční potenciál. Substituce nepropustných iontů sodíkem ruší akční potenciál. „Solitonový model“ nedokáže vysvětlit iontovou selektivitu ani reakce na změny napětí.
- Farmakologie
- Léčivý tetrodotoxin (TTX) blokuje akční potenciály při extrémně nízkých koncentracích. Bylo identifikováno místo působení TTX na sodíkový kanál.[34] Dendrotoxiny blokují draslíkové kanály. Tyto léky produkují kvantitativně předvídatelné změny akčního potenciálu.[33] „Solitonový model“ neposkytuje žádné vysvětlení těchto farmakologických účinků.
Akční vlny
Nedávný teoretický model, který navrhli Ahmed El Hady a Benjamin Machta, navrhuje, aby existovala mechanická povrchová vlna, která se šíří společně s elektrickým akčním potenciálem. Tyto povrchové vlny se nazývají „akční vlny“.[35] V modelu El Hady – Machta jsou tyto společně se šířící vlny poháněny změnami napětí na membráně způsobenými akčním potenciálem.
Viz také
Zdroje
- Federico Faraci (2013) "60. výročí Hodgkin-Huxleyova modelu: kritické hodnocení z historického a modelářského hlediska "
- Revathi Appali, Ursula van Rienen, Thomas Heimburg (2012) "Srovnání Hodgkin-Huxleyova modelu a teorie Soliton pro akční potenciál v nervech "
- Akční vlny v mozku, The Guardian, 1. května 2015.
- Ichiji Tasaki (1982) "Fyziologie a elektrochemie nervových vláken "
- Konrad Kaufman (1989) "Akční potenciály a elektrochemická vazba v makroskopické chirální fosfolipidové membráně ".
- Andersen, Jackson a Heimburg “Směrem k termodynamické teorii šíření nervových pulsů "
- Pradip Das; W.H. Schwarz (4. listopadu 1994). "Soliton v buněčné membráně". Fyzický přehled E. 51 (4): 3588–3612. Bibcode:1995PhRvE..51,3588D. doi:10.1103 / PhysRevE.51.3588. PMID 9963042.
- Přehodnocení mechaniky akčního potenciálu, Hodinky Princeton University Journal, 1. dubna 2015.
- Na (zvukové) stopě anestetik, Eurekalert, podle tiskové zprávy Kodaňská univerzita, 6. března 2007
- Kaare Græsbøll (2006). "Funkce nervů - působení anestetik" (PDF). Gama. 143. Základní úvod.
- Osamělé akustické vlny se šíří na rozhraní lipidové membrány, Phys.org 20. června 2014
Reference
- ^ Andersen, S; Jackson, A; Heimburg, T (2009). „Směrem k termodynamické teorii šíření nervových pulsů“ (PDF). Pokrok v neurobiologii. 88 (2): 104–113. doi:10.1016 / j.pneurobio.2009.03.002. PMID 19482227.
- ^ Hodgkin AL, Huxley AF, Katz B. (1952). „Proudy přenášené ionty sodíku a draslíku přes membránu obrovského axonu Loligo“. Fyziologický časopis. 116 (4): 424–448. doi:10.1113 / jphysiol.1952.sp004717. PMC 1392213. PMID 14946713.CS1 maint: více jmen: seznam autorů (odkaz)
Hodgkin AL, Huxley AF (1952). „Proudy nesené ionty sodíku a draslíku přes membránu obřího axonu Loligo“. Fyziologický časopis. 116 (4): 449–472. doi:10.1113 / jphysiol.1952.sp004717. PMC 1392213. PMID 14946713.
Hodgkin AL, Huxley AF (1952). „Složky membránové vodivosti v obřím axonu Loligo“. J. Physiol. 116 (4): 473–496. doi:10.1113 / jphysiol.1952.sp004718. PMC 1392209. PMID 14946714.
Hodgkin AL, Huxley AF (1952). „Duální účinek membránového potenciálu na vodivost sodíku v obřím axonu Loligo“. J. Physiol. 116 (4): 497–506. doi:10.1113 / jphysiol.1952.sp004719. PMC 1392212. PMID 14946715.
Hodgkin AL, Huxley AF (1952). „Kvantitativní popis membránového proudu a jeho aplikace na vedení a excitaci v nervu“. J. Physiol. 117 (4): 500–544. doi:10.1113 / jphysiol.1952.sp004764. PMC 1392413. PMID 12991237. - ^ Margineanu, D.-G; Schoffeniels, E. (1977). „Molekulární události a energetické změny během akčního potenciálu“. PNAS. 74 (9): 3810–3813. Bibcode:1977PNAS ... 74,3810M. doi:10.1073 / pnas.74.9.3810. PMC 431740. PMID 71734.
- ^ Hasenstaub, A; Callaway, E; Otte, S; Sejnowski, T (2010). „Metabolické náklady jako sjednocující princip řídící neuronovou biofyziku“. Sborník Národní akademie věd USA. 107 (27): 12329–12334. Bibcode:2010PNAS..10712329H. doi:10.1073 / pnas.0914886107. PMC 2901447. PMID 20616090.
- ^ Tasaki, Ichiji (13. října 1995). „Mechanické a tepelné změny v torpédovém elektrickém orgánu spojené s jeho postsynaptickými potenciály“. Sdělení o biochemickém a biofyzikálním výzkumu. 215 (2): 654–658. doi:10.1006 / bbrc.1995.2514. PMID 7488005.
- ^ Howarth, J V; Keynes, RD; Ritchie, J. M.; Muralt, A von (1. července 1975). „Produkce tepla spojená s průchodem jediného impulzu v pichavých čichových nervových vláknech“ (PDF). The Journal of Physiology. 249 (2): 349–368. doi:10.1113 / jphysiol.1975.sp011019. PMC 1309578. PMID 1236946.
- ^ Tasaki, I; Byrne, P. M. (1993). „Rychlá výroba tepla spojená s elektrickým buzením elektrických orgánů úhoře elektrického“. Biochem Biophys Res Commun. 197 (2): 910–915. doi:10.1006 / bbrc.1993.2565. PMID 8267630.
- ^ Tasaki, K; Kusano, K; Byrne, PM (1989). „Rychlé tepelné a mechanické změny v čichovém nervu garfishů spojené s rozšířeným impulzem“. Biophys J.. 55 (6): 1033–1040. Bibcode:1989BpJ .... 55.1033T. doi:10.1016 / s0006-3495 (89) 82902-9. PMC 1330571. PMID 2765644.
- ^ A b Howarth, J. V. (1975). "Produkce tepla v nemyelinovaných nervech". Filozofické transakce královské společnosti. 270 (908): 425–432. Bibcode:1975RSPTB.270..425H. doi:10.1098 / rstb.1975.0020. JSTOR 2417341. PMID 238239.
- ^ Tasaki, Ichiji (1982). Fyziologie a elektrochemie nervových vláken. Bethesda, Maryland: Academic Press Inc. (Londýn). ISBN 978-0-12-683780-3.
- ^ Kaufmann, Konrad (1989). Akční potenciály a elektrochemická vazba v makroskopické chirální fosfolipidové membráně. Caruaru, Brazílie.
- ^ Xin-Yi, Wang (1985). "Osamělá vlna a nerovnovážný fázový přechod v kapalných krystalech". Fyzický přehled A. 32 (5): 3126–3129. Bibcode:1985PhRvA..32.3126X. doi:10.1103 / PhysRevA.32.3126. PMID 9896466.
- ^ Heimburg, T., Jackson, A.D. (12. července 2005). „O šíření solitonů v biomembránách a nervech“. Proc. Natl. Acad. Sci. USA. 102 (2): 9790–9795. Bibcode:2005PNAS..102.9790H. doi:10.1073 / pnas.0503823102. PMC 1175000. PMID 15994235.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Heimburg, T., Jackson, A.D. (2007). „O akčním potenciálu jako pulzu šířící se hustoty a roli anestetik“. Biophys. Rev. Lett. 2: 57–78. arXiv:fyzika / 0610117. Bibcode:2006fyzika..10117H. doi:10.1142 / S179304800700043X.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Andersen, S.S.L., Jackson, A.D., Heimburg, T. (2009). "Směrem k termodynamické teorii šíření nervových pulsů". Prog. Neurobiol. 88 (2): 104–113. doi:10.1016 / j.pneurobio.2009.03.002. PMID 19482227.CS1 maint: více jmen: seznam autorů (odkaz)[mrtvý odkaz ]
- ^ Griesbauer, J; Bossinger, S; Wixforth, A; Schneider, M (9. května 2012). „Propagace 2D tlakových pulzů v lipidových monovrstvách a její možné důsledky pro biologii“. Dopisy o fyzické kontrole. 108 (19): 198103. arXiv:1211.4104. Bibcode:2012PhRvL.108s8103G. doi:10.1103 / PhysRevLett.108.198103. PMID 23003093.
- ^ A b Shrivastava, Shamit; Schneider, Matthias (18. června 2014). „Důkazy pro dvourozměrné osamělé zvukové vlny v rozhraní řízeném lipidy a jeho důsledky pro biologickou signalizaci“. Journal of the Royal Society Interface. 11 (97): 20140098. doi:10.1098 / rsif.2014.0098. PMC 4078894. PMID 24942845.
- ^ Griesbauer, J; Bossinger, S; Wixforth, A; Schneider, M (19. prosince 2012). "Současné šíření napěťových a tlakových pulzů v lipidových monovrstvách vepřového mozku a syntetických lipidů". Fyzický přehled E. 86 (6): 061909. arXiv:1211.4105. Bibcode:2012PhRvE..86f1909G. doi:10.1103 / PhysRevE.86.061909. PMID 23367978.
- ^ Shrivastava, Shamit (leden 2014). NELINEÁRNÍ SOLITÁRNÍ ZVUKOVÉ VLNY V LIPIDOVÝCH MEMBRÁNÁCH A JEJICH MOŽNÁ ROLE V BIOLOGICKÉM SIGNALIZACI (1. vyd.). Boston, MA 02215 USA: Thesis, Boston University.CS1 maint: umístění (odkaz)
- ^ Abbott, B.C., Hill, A.V., Howarth, J.V. (1958). "Pozitivní a negativní teplo spojené s nervovým impulsem". Sborník Královské společnosti B. 148 (931): 149–187. Bibcode:1958RSPSB.148..149A. doi:10.1098 / rspb.1958.0012. PMID 13518134.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Iwasa, K., Tasaki I., Gibbons, R. (1980). "Otok nervových vláken spojený s akčními potenciály". Věda. 210 (4467): 338–9. Bibcode:1980Sci ... 210..338I. doi:10.1126 / science.7423196. PMID 7423196.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Griesbauer, J; Wixforth, A; Schneider, MF (15. listopadu 2009). „Šíření vln v lipidových monovrstvách“. Biofyzikální deník. 97 (10): 2710–2716. Bibcode:2009BpJ .... 97,2710G. doi:10.1016 / j.bpj.2009.07.049. PMC 2776282. PMID 19917224.
- ^ Villagran Vargas, E., Ludu, A., Hustert, R., Gumrich, P., Jackson, A.D., Heimburg, T. (2011). "Periodická řešení a refrakterní periody v teorii solitonů pro nervy a akátový femorální nerv". Biofyzikální chemie. 153 (2–3): 159–167. arXiv:1006.3281. doi:10.1016 / j.bpc.2010.11.001. PMID 21177017.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Heimburg, T. (2010). "Lipidové iontové kanály". Biophys. Chem. 150 (1–3): 2–22. arXiv:1001.2524. Bibcode:2010arXiv1001.2524H. doi:10.1016 / j.bpc.2010.02.018. PMID 20385440.
- ^ Heimburg, T., Jackson, A.D. (2007). „Termodynamika celkové anestezie“. Biophys. J. 92 (9): 3159–65. arXiv:fyzika / 0610147. Bibcode:2007BpJ .... 92,3159H. doi:10.1529 / biophysj.106.099754. PMC 1852341. PMID 17293400.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Rall, W a Shepherd, GM (1968) Teoretické rekonstrukce dendrodendritických synaptických interakcí v čichové baňce. J. Neurophysiol 31, 884-915.http://jn.physiology.org/content/jn/31/6/884.full.pdf
- ^ Hodgkin; Katz (1949). „Vliv teploty na elektrickou aktivitu obřího axonu chobotnice“. J. Physiol. 109 (1–2): 240–249. doi:10.1113 / jphysiol.1949.sp004388. PMC 1392577. PMID 15394322.
- ^ Tasaki, Ichiji (1949). "Srážka dvou nervových impulsů v nervovém vlákně". Biochim Biophys Acta. 3: 494–497. doi:10.1016/0006-3002(49)90121-3.
- ^ Gonzalez, Alfredo; Budvytyte, Rima; Mosgaard, Lars D; Nissen, Søren; Heimburg, Thomas (10. září 2014). „Průnik akčních potenciálů během kolize ve středních a laterálních obřích axonech bezobratlých“. Fyzická kontrola X. 4 (3): 031047. arXiv:1404.3643. Bibcode:2014PhRvX ... 4c1047G. doi:10.1103 / PhysRevX.4.031047.
- ^ Sander, HW, Collision Testing in J Kimura Peripheral Nerve Diseases https://books.google.com/books?id=jp05zU9vxo8C&pg=PA359&lpg=PA359&dq=collision+test+neurophysiology&source=bl&ots=PTz3H5Mn-t&sig=LUtPKvs1ad8q0wX8zIQ712mNG7E&hl=en&sa=X&ei=dE9iVfvlENOMyASFz4OQAg&ved=0CB0Q6AEwADgK#v=onepage&q=collision%20test% 20neurofyziologie & f = nepravda
- ^ Eckl, C; Mayer, AP; Kovalev, AS (3. srpna 1998). "Existují povrchové akustické solitony?". Dopisy o fyzické kontrole. 81 (5): 983–986. Bibcode:1998PhRvL..81..983E. doi:10.1103 / PhysRevLett.81.983.
- ^ Shrivastava, Shamit; Kang, Kevin; Schneider, Matthias F (30. ledna 2015). „Osamělé rázové vlny a adiabatický fázový přechod v lipidových rozhraních a nervech“. Fyzický přehled E. 91 (12715): 012715. arXiv:1411.2454. Bibcode:2015PhRvE..91a2715S. doi:10.1103 / PhysRevE.91.012715. PMID 25679650.
- ^ A b Hille, Bertil (2001). Iontové kanály excitovatelných membrán (3. vyd.). Sunderland, Massachusetts: Sinauer. ISBN 9780878933211.
- ^ Catterall, WA (2014). "Struktura a funkce napěťově řízených sodíkových kanálů při atomovém rozlišení". Experimentální fyziologie. 99 (1): 35–51. doi:10.1113 / expphysiol.2013.071969. PMC 3885250. PMID 24097157.
- ^ El Hady, A., Machta, B. (2015). "Mechanické povrchové vlny doprovázejí šíření akčního potenciálu". Příroda komunikace. 6: 6697. arXiv:1407.7600. Bibcode:2015NatCo ... 6.6697E. doi:10.1038 / ncomms7697. PMID 25819404.CS1 maint: více jmen: seznam autorů (odkaz)