Ornithosuchidae - Ornithosuchidae
| Ornithosuchidae | |
|---|---|
 | |
| Obnova života Ornithosuchus úklid na rhynchosaur Hyperodapedon | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Třída: | Reptilia |
| Clade: | Pseudosuchia |
| Rodina: | †Ornithosuchidae von Huene, 1908 |
| Zadejte druh | |
| †Ornithosuchus woodwardi Newton, 1894 | |
| Rody | |
†Aenigmaspina ? | |
Ornithosuchidae je vyhynulý rodina z pseudosuchian archosaury (vzdálení příbuzní moderní krokodýli ) z Trias doba. Ornitosuchidi byli čtyřnohý a fakultativně bipedal, což znamená, že měli schopnost chodit na dvou nohách po krátkou dobu. Měli výrazné pokleslé čenichy, jedinečné kotníkové kosti s „obráceným krokodýlem“ a několik dalších funkcí, které paleontologové používají k jejich odlišení od ostatních archosaurů. Ornitosuchidy byly během roku geograficky rozšířené Karnian a Norian etapy Pozdní trias, se členy známými z Argentina, Brazílie a Spojené království. Čtyři rody, Ornithosuchus, Venaticosuchus, Dynamosuchus,[1] a Riojasuchus jsou v současnosti známé.[2] Rodina byla poprvé postavena německým paleontologem Friedrich von Huene v roce 1908.[3]
Popis
Lebka
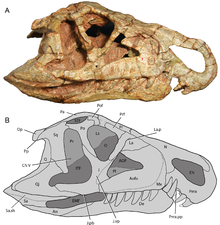
Ornithosuchids lze identifikovat podle přítomnosti klenuté diastema, mezera mezi zuby v přední části čenichu. Když je čelist zavřená, dvě velké zakřivené zubař (dolní čelist) zuby zapadají do diastému, který je umístěn mezi premaxilla a horní čelist. Na stěně diastému jsou dvě mělké prohlubně, aby se do nich mohly umístit tyto zuby. Velké zuby zubu Ornithosuchus a Riojasuchus jsou umístěny za menším zubním zubem, který vyčnívá z čelisti. Tento typ polohy zubu není vidět u žádných jiných bazálních archosaurů. Dalším charakteristickým rysem ornithosuchidů je jejich neobvyklý pokleslý, převislý čenich, který je vidět na Riojasuchus a Venaticosuchus, ale ne Ornithosuchus.[4]
Několik dalších funkcí odlišuje ornitosuchidy od všech ostatních raných archosaurů. Ornithosuchus a Riojasuchus oba mají malý fenestr nebo otvor mezi palatine a pterygoidní kosti z patro, tj. střecha úst. Kontakt mezi nosní a prefrontální kosti lebky je malá nebo chybí, je vyloučena velkým kontaktem mezi čelní strany a slzy. V jiných archosaurech, včetně rauisuchians, aetosaurs, ptakoještěr, a dinosauromorfy, nosní-prefrontální kontakt odděluje frontální od slzného.[4]
Postkraniální kostra
Postkraniální kostra je téměř úplně známa Riojasuchus, neúplné v Ornithosuchus, a zcela neznámý v Venaticosuchus. Výsledkem je, že není jisté, zda se všechny postkraniální rysy, které se zdají být jedinečné pro ornitosuchidy, skutečně vyskytly u všech členů rodiny. Ornitosuchidy známé ze slušných postkraniálních pozůstatků měly obvykle asi 9 krční (krk), 14-15 hřbetní (vzadu), 3 křížový (kyčel) a více než 20 kaudálních (ocas) obratel. Nad každým obratlem byl pár kostnatých scutů známých jako osteodermy.[2]
The stehenní kost (stehenní kost) má výrazný přední trochanter. Přední trochanter, někdy známý jako "menší trochanter" (ale nesouvisí s menší trochanter stehenní kosti u lidí), je vyvýšenina na vnějším povrchu stehenní kosti, v blízkosti hlavice stehenní kosti. Pravděpodobně to byl kurzor pro iliofemoralis cranialis sval, který pomáhá zvedat nohu. Většina archosaurů a příbuzných archosaurů postrádá zřetelný přední trochanter, ale ornitosuchidy jsou výjimkou, spolu s většinou dinosauromorfy (dinosauři a jejich blízcí příbuzní).[5]
Stejně jako stehenní kost fibula (vnější holenní kost) má také výrazný bod pro zavedení svalů. Dotyčný sval je iliofibularis, což pomáhá napínat končetiny. Ve většině archosaurů je iliofibularis se zavádí na lýtkovou část pomocí malého hřebene na proximální části lýtkové kosti, blízko kolena. Ornithosuchidové však mají mnohem větší knoflíkovitý tvar iliofibularis zaváděcí bod umístěný přibližně v polovině dříku lýtkové kosti. Fytosaury a aetosaury také sdílejí připevňovací bod podobný knoflíku uprostřed lýtkové kosti, takže není jasné, zda je případ ornithosuchidů jedinečným případem konvergentní evoluce, nebo alternativně zachování rysu nezávisle ztraceného několika liniemi archosaurů.[6]
Na rozdíl od většiny ostatních raných archosaurů, pedálu homosexuálové (nejvzdálenější kosti chodidel, které tvoří drápy) jsou bočně stlačeny. Jsou ostré a zakřivené. Unguálové jsou velmi hlubokí, jsou vyšší než delší, zejména na vnitřních číslicích. Tento typ drápu není vidět u žádného jiného triasového archosaura kromě pterosaurů.[4]

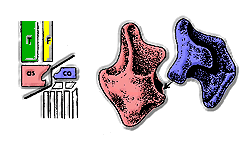
Hlavní skupiny archosaurů se často navzájem odlišují na základě struktury kotníků. Ve většině crurotarsans, astragalus má konvexní projekci, která zapadá do konkávního prostoru v patní kost. Tento stav je často označován jako „krokodýlí normální“ kotník, protože je nejběžnějším typem kotníku u crurotarsanů. Ornithosuchidové jsou mezi crurotarsany a všemi ostatními archosaury jedineční, protože mají kotník „obrácený krokodýlem“. V kotníku s „krokodýlem obráceným“ je umístění konkávnosti obráceno: místo toho, aby byl na kalkanea, je na astragalu. V ornithosuchidech má kalkanea konvexní projekci, která je analogická konvexní projekci na „krokodýlí normální“ astragalus.[4]
Fylogeneze
Ornithosuchidae se obecně považuje za větší clade Suchia, což zahrnuje aetosaury, rauisuchiany a crocodylomorfy.[7][8][9] Níže je cladogram založený na Nesbittovi (2011), který ukazuje umístění Ornithosuchidae v Archosauriformes.[6]
| Archosauriformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Reference
- ^ A b Müller, Rodrigo T .; Von Baczko, M. Belén; Desojo, Julia B .; Nesbitt, Sterling J. (31. ledna 2020). „První ornitosuchid z Brazílie a jeho makroevoluční a fylogenetické důsledky pro fauny pozdního triasu v Gondwaně“ (PDF). Acta Palaeontologica Polonica. 65. doi:10.4202 / app.00652.2019.
- ^ A b Baczko, M. Belén von; Ezcurra, Martín D. (01.01.2013). „Ornithosuchidae: skupina triasových archosaurů s jedinečným kotníkovým kloubem“. Geologická společnost, Londýn, speciální publikace. 379 (1): 187–202. doi:10.1144 / SP379.4. ISSN 0305-8719.
- ^ Huene, F. von. 1908. „Die Dinosaurier der europäischen Triasformation mit Berücksichtigung der aussereuropäischen Vorkommnisse“. Geologische und Paläontologische Abhandlungen 1(Suppl.): 1–419
- ^ A b C d Sereno, P.C. (1991). „Bazální archosaury: fylogenetické vztahy a funkční důsledky“. Časopis paleontologie obratlovců. 11 (Příloha 4): 1–53. doi:10.1080/02724634.1991.10011426.
- ^ Langer, Max. Benton, Michael J. (6. listopadu 2006). „Raní dinosauři: fylogenetická studie“ (PDF). Journal of Systematic Palaeontology. 4 (4): 309–358. doi:10.1017 / s1477201906001970. ISSN 1477-2019.
- ^ A b Nesbitt, S.J. (2011). „Počáteční vývoj archosaurů: vztahy a původ hlavních subtypů“ (PDF). Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112.
- ^ Nesbitt, S.J .; Norrell, M.A. (2006). „Extrémní konvergence v plánech těla raného sucha (Archosauria) a ornithomimidních dinosaurů (Theropoda)“. Sborník Královské společnosti B. 273 (1590): 1045–1048. doi:10.1098 / rspb.2005.3426. PMC 1560254. PMID 16600879.
- ^ Nesbitt, S.J. (2007). "Anatomie Effigia okeeffeae (Archosauria, Suchia), sbližování podobné teropodům a distribuce souvisejících taxonů “ (PDF). Bulletin of the American Museum of Natural History. 302: 1–84. doi:10.1206 / 0003-0090 (2007) 302 [1: TAOEOA] 2.0.CO; 2. hdl:2246/5840.
- ^ Brusatte, S.L .; Benton, M. J.; Desojo, J.B .; Langer, M.C. (2010). „Fylogeneze vyšší úrovně Archosaurie (Tetrapoda: Diapsida)“ (PDF). Journal of Systematic Palaeontology. 8 (1): 3–47. doi:10.1080/14772010903537732.
externí odkazy
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||

