Mesosaurs („střední ještěrky“) byla skupina malých vodních plazů, kteří žili na počátku Permu období zhruba Před 299 až 270 miliony let. Mesosaurs byli první známí vodní plazi poté, co se zjevně vrátil k vodnímu životnímu stylu od více suchozemských předků. Přesné, jak se předkové pozemských mezosaurů stali, zůstává nejisté; nedávný výzkum nemůže s jistotou určit, jestli první amniotes byli zcela pozemští nebo jen obojživelní.[1] Většina autorů považuje mesosaury za vodní,[1][2] ačkoli dospělá zvířata mohla být spíše obojživelní než zcela vodní, což naznačuje jejich mírné přizpůsobení kostry semiakvatickému životnímu stylu.[3][4] Podobně jsou jejich příbuznosti nejisté; mohou být buď nejzákladnější sauropsidy,[5][6] nebo mezi nejzákladnější parareptilové.[7][8]
Fylogenetická poloha mezosaurů má důležitý vliv na definici Reptilie. V jedné z prvních velkých fylogenetických studií amniotů (obratlovců kladoucích vajíčka na pevninu) Gauthier et al. (1988) umístili Mesosauridae do skupiny zvané Parareptilia.[10] Parareptilia znamená „na straně plazů“ a byla umístěna venku clade Reptilia, která byla považována za skupina korun. Jako korunní skupina zahrnovala Reptilia poslední společný předek dvou hlavních linií živých plazů -diapsidy a želvy - a všichni potomci tohoto společného předka. V tomto smyslu byla Reptilia a uzlový taxon protože první společný předek plazů by byl „uzlem“ na fylogenetickém stromu. Podle této fylogeneze bylo mnoho vyhynulých forem tradičně považovaných za plazy, včetně mezosaurů, vyloučeno ze skupiny, protože byly mimo uzel.[9]
Fosílie jihoamerického mezosaura
Studie Laurina a Reisze (1995) byla druhou významnou fylogenetickou analýzou amniotů.[11] Jako Gauthier et al.Laurin a Reisz použili Reptilii jako skupinu korun a umístili mesosaury mimo skupinu. Jejich fylogeneze se lišila v tom, že Gauthierovi parareptili et al. byli nyní považováni za blízké příbuzné želv v rámci korunní skupiny Reptilia. Laurin a Reisz přijali jméno Sauropsida jako uzlový taxon, včetně posledního společného předka mesosaurů a Reptilií. Amnioti se tradičně dělí do dvou skupin: tzv. Linie savců Synapsida a linie plazů zvaná Reptilia nebo Sauropsida. Ve skutečnosti studie Gauthiera (1994) definovala Sauropsidu jako všechny amnioty, které se více vztahovaly k plazům než k savcům, což znamenalo, že Sauropsida byla kmenový taxon zahrnující celou plazovou linii nebo plazivý „kmen“ Amniota (Synapsida byl kmen savců). Podle této fylogeneze je jedinou skupinou, která brání tomu, aby Sauropsida byla rovnocenná s Reptilií, jsou mesosaurové.[9]
Novější fylogenetické analýzy, jako například Modesto (1999), podporují Gauthier et al. (1988) umístěním mezosaurů na parareptily.[7] Tyto fylogeneze však následují Laurina a Reisze (1995) při umisťování Parareptilii do korunní skupiny Reptilia, což znamená, že mezosaury jsou opět členy Reptilie. Pomocí Laurinovy a Reiszovy uzlové definice Sauropsidy jako „posledního společného předka mesosaurů, testudinů a diapsidů a všech jeho potomků“,[11] Sauropsida a Reptilia jsou ekvivalentní seskupení; mesosaury a testudiny jsou si navzájem bližší, než jakákoli skupina má diapsidy,[A] což znamená, že clade obsahující testudiny a diapsidy (což by byla korunní skupina Reptilia) musí také obsahovat mesosaury. Vzhledem k tomu, že Reptilia byla pojmenována dříve než Sauropsida, používá se nejčastěji v moderních fylogenetických analýzách.[9]
Fylogenetická analýza z roku 2017, kterou provedli Laurin (který dříve publikoval studii z roku 1995) a Piñeiro obnovili mezosaury jako bazální člen Sauropsida / Reptilia a již nejsou přítomni v Parareptilii, přičemž Parareptilia byla nově definována jako zahrnující bývalé členy Procolophonomorpha (zjištěno, že je parafyletický), Millerosauria, Pareiasauria, a Pantestudiny, přičemž poslední dva byly shledány jako sesterské skupiny navzájem. Bylo také zjištěno, že Parareptilia skutečně hnízdila uvnitř Diapsidy jako sesterská skupina Neodiapsida.[13]
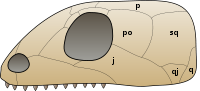
V roce 2012 se ukázalo, že Mesosaurus má otvory v zadní části lebky zvané nižší temporální fenestry, charakteristika, o které se kdysi myslelo, že je přítomna pouze v synapsidech a diapsidech.[14] Stav v lebce Mesosaurus je nejvíce podobný tomu u synapsidových lebek, protože oběma chybí horní temporální fenestry diapsidů. Nižší temporální fenestry jsou zatím známy pouze v Mesosaurus, ale mohou být přítomni ve všech mesosaurech. Přítomnost nebo nepřítomnost časových fenester je důležitým faktorem ve fylogenezi mesosaurů a dalších amniotů, protože tři hlavní skupiny amniotů - Synapsida, Diapsida a Anapsida - byly pojmenovány podle počtu otvorů v jejich lebce; Diapsida znamená „dva oblouky“ v odkazu na dva pruhy, které uzavírají horní a dolní fenestr, Synapsida znamená „tavený oblouk“ v odkazu na jeden pruh v dolní části lebky uzavírající jeden fenestr a Anapsida znamená „žádný oblouk „v odkazu na lebky, které postrádají tyče nebo fenestry. Mesosaurové byli tradičně klasifikováni jako anapsidové, protože se předpokládalo, že jim chyběly časové fenestry. Výskyt fenestra v amniotech byl však ve skupině uznáván jako velmi variabilní rys po mnoho let před jejich objevením v Mesosaurus; mnoho anapsidů jako např Candelaria, Bolosaurus, a lanthanosuchoidy mají nižší temporální fenestry. Kromě toho je Anapsida v posledních fylogenetických analýzách zřídka považována za platný clade.
Lebka zobecněné anapsidy.
Lebka zobecněného synapsidu.
Fylogenetická poloha mezosaurů ovlivňuje současné chápání toho, jak se u amniotů vyvinula temporální fenestra. Pokud je fylogeneze produkovaná Laurinem a Reiszem (1995) správná v tom, že mesosaury jsou bazální sauropsidy, dolní temporální fenestra může být u amniotů primitivním rysem, který je přítomen u posledního společného předka amniota. Synapsidy by si zachovaly svůj fenestr, stejně jako sauropsidy, kromě želv a většiny parareptilů. Další možností podle Laurinovy a Reiszovy fylogeneze je to, že dolní temporální fenestry se vyvíjely nezávisle u mesosaurů, synapsidů, diapsidů a některých parareptilů a že nedostatek fenestra je u amniotů primitivní funkcí. Pokud jsou místo toho mesosaurové členy Parareptilia, není přítomnost časových fenester pravděpodobně u amniotů primitivním rysem a dolní temporální fenestry u mesosaurů mohou být charakteristické pro linii bazálních parareptilů, která zahrnuje také lanthanosuchoidy nesoucí fenestru a Bolosaurus.[14]
Biologie
Dlouho se o nich myslelo, že jsou to pobřežní formy, které pravděpodobně obývají relativně mělkou vodu,[15] ale nedávný výzkum naznačuje, že přinejmenším ti z Uruguaye obývali spíše hypersalinní prostředí než pobřežní mořské prostředí.[16] Nedávno popsaná embrya ukazují, že pachyostóza žeber (která byla silnější a hustší než u suchozemských tetrapodů) se vyvinula ještě před vylíhnutím, což naznačuje, že mezosaurové byli schopni plavat při narození nebo krátce poté. Zjevně to nebyli příliš rychlí plavci s optimální rychlostí plavání odhadovanou na 0,15 až 0,86 m / s, ale muselo to být o něco rychlejší než rychlost jejich hlavní kořisti, pygocephalomorf korýši.[17] Jejich reprodukční režim je poněkud nejistý, protože asociace mezi dospělými a možnými embryi in utero naznačuje viviparitu, stejně jako u mnoha vodních plazů, ale bylo také nalezeno potenciálně izolované vejce.[18]
Nedávno důkazy o predaci u obou korýšů pygocephalomorfu a příslušníci jejich vlastního druhu byla stanovena. Předpokládá se, že mesosaury byly obecně přizpůsobeny hypersalinním stanovištím.[19]
Studie proporcí trupu a ocasu páteře Mesosaurus navrhuje, že zatímco mladiství mohou být zcela vodní, dospělí možná strávili nějaký čas na souši; toto je dále potvrzeno vzácností dospělých zvířat ve vodním prostředí a některými výkaly vykazujícími známky sušící zlomeniny. Je však těžké říci, jak suchozemští byli, protože stejná studie uvádí, že pozemské shánění potravy by bylo obtížné vzhledem k jejich zvláštnostem pro vodní život.[20]
Poznámky
^ Studie využívají molekulární fylogenetika, které zkoumají geny a proteiny živých organismů, naznačují, že testudiny (želvy) jsou diapsidy. Tyto studie ukazují, že mesosaurové netvoří clade s želvami, které vylučují diapsidy, ale fosilní důkazy stále naznačují, že mesosaurové tvoří skupinu s parareptily. Ve většině nedávných studií se Reptilia nepoužívá jako skupina korun a stále obsahuje mezosaury a parareptilii.
Reference
^ AbCanoville, A. a M. Laurin. 2010. Vývoj humerální mikroanatomie a životního stylu u amniotů a některé komentáře k paleobiologickým závěrům. Biological Journal of the Linnean Society 100: 384–406.
^Modesto, S. 2006. Lebeční kostra raně permského vodního plaza Mesosaurus tenuidens: důsledky pro vztahy a paleobiologii. Zoological Journal of the Linnean Society 146: 345-368.
^Piñeiro, G. 2008. Los mesosaurios y otros fosiles de fines del Paleozoico; str. 179–205 v publikaci D. Perera (ed.), Fósiles de Uruguay. DIRAC, Montevideo.
^Pablo Nuñez Demarco a kol. Byl Mesosaurus plně vodním plazem? Přední. Ecol. Evol, publikováno online 27. července 2018; doi: 10.3389 / fevo.2018.00109
^Laurin, M. a R. R. Reisz. 1995. Přehodnocení časné fylogeneze amniote. Zoological Journal of the Linnean Society 113: 165-223.
^ AbPiñeiro, G .; Ferigolo, J .; Ramos, A .; Laurin, M. (2012). „Kraniální morfologie raně permského mesosauridu Mesosaurus tenuidens a přehodnocen vývoj dolní časové fenestrace“. Comptes Rendus Palevol. 11 (5): 379–391. doi:10.1016 / j.crpv.2012.02.001.
^Oelofsen, B. a D. C. Araújo. 1983. Paleoekologické důsledky distribuce plazů mesosauridů v permském moři Irati (povodí Paraná), Jižní Amerika. Revista Brasileira de Geociências 13:1–6.
^Villamil, J. N .; Demarco, P. N .; Meneghel, M .; Blanco, R.E .; Jones, W .; Rinderknecht, A. S .; Laurin, M .; Pineiro, G. (2015). „Optimální odhady rychlosti plavání v raně permském mesosauridu Mesosaurus tenuidens (Gervais 1867) z Uruguaye “. Historická biologie. 28 (7): 963–971. doi:10.1080/08912963.2015.1075018. S2CID85846750.
^Piñeiro, G .; Ferigolo, J .; Meneghel, M .; Laurin, M. (2012). "Nejstarší známá plodová embrya naznačují viviparitu u mesosaurů". Historická biologie. 24 (6): 620–630. doi:10.1080/08912963.2012.662230. S2CID59475679.
^Rivaldo R. Da Silva; Jorge Ferigolo; Piotr Bajdek; Graciela H. Piñeiro (2017). "Stravovací návyky Mesosauridae". Hranice ve vědě o Zemi. 5: Článek 23. doi: 10.3389 / feart.2017.00023.
^Pablo Nuñez Demarco a kol. Byl Mesosaurus plně vodním plazem? Přední. Ecol. Evol, publikováno online 27. července 2018; doi: 10.3389 / fevo.2018.00109