Syntetický biologický obvod - Synthetic biological circuit
| Část série článků o |
| Syntetická biologie |
|---|
| Syntetické biologické obvody |
| Úpravy genomu |
| Umělé buňky |
| Xenobiologie |
| Další témata |

1: RNA polymeráza, 2: Represor, 3: Promotér, 4: Operátor, 5: Laktóza, 6: lacZ, 7: krajkový, 8: lacA.
Syntetické biologické obvody jsou aplikací syntetická biologie kde biologické části uvnitř a buňka jsou navrženy k provádění logických funkcí napodobujících funkce pozorované v elektronické obvody. Rozsah aplikací sahá od jednoduchého vyvolání výroby až po přidání měřitelného prvku GFP, na existující přirozený biologický obvod, k implementaci zcela nových systémů mnoha částí.[1]
Cílem syntetické biologie je generovat řadu laditelných a charakterizovaných částí nebo modulů, pomocí kterých lze snadno navrhnout a implementovat jakýkoli požadovaný syntetický biologický obvod.[2] Tyto obvody mohou sloužit jako metoda pro úpravu buněčných funkcí, vytváření buněčných odpovědí na podmínky prostředí nebo ovlivnění buněčného vývoje. Implementací racionálních, kontrolovatelných logických prvků v celulárních systémech mohou vědci využívat živé systémy tak, jak byly navrženy “biologické stroje "provádět celou řadu užitečných funkcí.[1]
Dějiny
První obvod přírodních genů podrobně studovaný byl lac operon. Ve studiích diauxický růst z E-coli na médiu se dvěma cukry, Jacques Monod a Francois Jacob objevil to E-coli přednostně spotřebovává snadněji zpracované glukóza před přepnutím na laktóza metabolismus. Zjistili, že mechanismus, který řídil metabolickou „přepínací“ funkci, byl dvoudílný kontrolní mechanismus na lac operonu. Pokud je v buňce přítomna laktóza, enzym β-galaktosidáza se vyrábí k přeměně laktózy na glukóza nebo galaktóza. Pokud laktóza v buňce chybí, potlačuje lac represor produkci enzymu β-galaktosidázy, aby zabránil neúčinným procesům v buňce.
Lac operon se používá v biotechnologie průmysl pro výrobu rekombinantní bílkoviny pro terapeutické použití. Gen nebo geny pro produkci exogenní bílkoviny jsou umístěny na a plazmid pod kontrolou lac promotoru. Zpočátku jsou buňky pěstovány v médiu, které neobsahuje laktózu nebo jiné cukry, takže nové geny nejsou exprimovány. Jakmile buňky dosáhnou určitého bodu svého růstu, Isopropyl β-D-1-thiogalaktopyranosid (IPTG) je přidáno. IPTG, molekula podobná laktóze, ale se sírovou vazbou, která není hydrolyzovatelná, takže ji E. Coli nestráví, se používá k aktivaci nebo "vyvolat "produkce nového proteinu. Jakmile jsou buňky indukovány, je obtížné odstranit IPTG z buněk, a proto je obtížné zastavit expresi."
Dva rané příklady syntetických biologických obvodů byly publikovány v Příroda v roce 2000. Jeden, Tim Gardner, Charles Cantor a Jim Collins pracovat v Bostonská univerzita, předvedl „bistabilní“ přepínač E-coli. Přepínač se zapne zahřátím bakteriální kultury a vypne se přidáním IPTG. Použili GFP jako reportér pro svůj systém.[3] Druhý tím, že Michael Elowitz a Stanislas Leibler, ukázal, že tři represorové geny mohou být spojeny za vzniku smyčky negativní zpětné vazby nazývané Represor který produkuje soběstačné oscilace hladin bílkovin v E-coli.[4]
V současné době jsou syntetické obvody rychle se rozvíjející oblastí výzkumu v biologie systémů s více publikacemi, které podrobně popisují syntetické biologické obvody, každý rok.[5] Významný zájem byl také o podporu vzdělávání a dosah: Mezinárodní soutěž v geneticky upravených strojích[6] spravuje tvorbu a standardizaci Bio cihla části jako prostředek umožňující vysokoškolákům a středoškolákům navrhovat vlastní syntetické biologické obvody.
Zájem a cíle
Pro použití syntetických biologických obvodů existují okamžité i dlouhodobé aplikace, včetně různých aplikací pro metabolické inženýrství, a syntetická biologie. Mezi úspěšně prokázané patří farmaceutická výroba,[7] a výroba paliva.[8] Metody zahrnující přímé genetické zavedení však nejsou ze své podstaty účinné bez uplatnění základních principů syntetických buněčných obvodů. Například každý z těchto úspěšných systémů využívá metodu k zavedení indukce nebo výrazu vše-nebo-žádný. Jedná se o biologický obvod, kde je jednoduchý represor nebo promotér se zavádí s cílem usnadnit vytvoření produktu nebo inhibici konkurenční cesty. S omezeným porozuměním celulárním sítím a přirozeným obvodům je však bráněno implementaci robustnějších schémat s přesnější kontrolou a zpětnou vazbou. V tom spočívá okamžitý zájem o syntetické buněčné obvody.
Vývoj v porozumění buněčným obvodům může vést k vzrušujícím novým modifikacím, jako jsou buňky, které mohou reagovat na podněty prostředí. Mohly by být například vyvinuty buňky, které signalizují toxické prostředí a reagují aktivací drah používaných k degradaci vnímaného toxinu.[9] K vývoji takové buňky je nutné vytvořit složitý syntetický buněčný obvod, který dokáže odpovídajícím způsobem reagovat na daný podnět.
Vzhledem k tomu, že syntetické buněčné obvody představují formu řízení buněčných aktivit, lze usuzovat, že při úplném porozumění buněčným cestám „plug and play“[1] buňky s dobře definovanými genetickými obvody mohou být konstruovány. Obecně se věří, že pokud se vygeneruje správný soubor nástrojů pro součásti,[10] lze vyvinout syntetické buňky, které implementují pouze cesty nezbytné pro reprodukci přežití buněk. Z této buňky lze považovat za minimální genom buňka, lze přidat kousky ze sady nástrojů a vytvořit dobře definovanou cestu s příslušnými syntetickými obvody pro efektivní zpětnovazební systém. Kvůli základní konstrukční metodě uzemnění a navrhované databázi mapovaných částí obvodů lze k přepracování buněk a modelování buněk pro snadné řešení problémů a prediktivní chování a výnosy použít techniky zrcadlení těch, které se používají k modelování počítačových nebo elektronických obvodů.
Ukázkové obvody
Oscilátory
- Represor
- Nastavitelný syntetický oscilátor pro savce
- Bakteriálně laditelný syntetický oscilátor
- Spojený bakteriální oscilátor
- Globálně spojený bakteriální oscilátor
Elowitz a kol. a Fung a kol. vytvořil oscilační obvody, které používají několik samoregulačních mechanismů k vytvoření časově závislé oscilace exprese genového produktu.[11][12]
Bistabilní spínače
- Přepnout spínač
Gardner a kol. používá vzájemnou represi mezi dvěma řídicími jednotkami k vytvoření implementace přepínacího přepínače schopného řídit buňky bistabilním způsobem: přechodné podněty vedoucí k trvalým reakcím[3].
Logické operátory

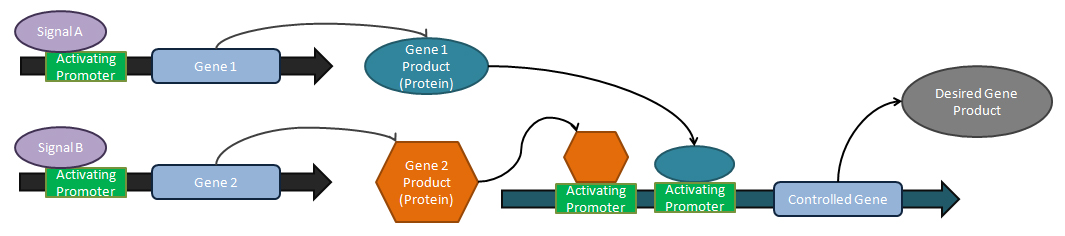

Analogové tunery
Pomocí negativní zpětné vazby a identických promotorů mohou obvody genů linearizátoru ukládat jednotnou genovou expresi, která lineárně závisí na koncentraci extracelulárního chemického induktoru.[15]
Regulátory heterogenity genové exprese
Syntetické genové obvody mohou řídit genovou expresi heterogenitu lze řídit nezávisle na střední hodnotě genové exprese.[16]
Jiné upravené systémy
Inženýrské systémy jsou výsledkem implementace kombinací různých řídicích mechanismů. Mechanismus omezeného počítání byl implementován pulzní řízenou genovou kaskádou[17] a aplikace logických prvků umožňuje genetické „programování“ buněk jako ve výzkumu Tabora a kol., který syntetizoval fotocitlivý program detekce bakteriálních hran.[18]
Návrh obvodu

Poslední vývoj v roce 2006 umělá genová syntéza a odpovídající zvýšení konkurence v tomto odvětví vedlo k významnému poklesu ceny a čekací doby genové syntézy a pomohlo zlepšit metody používané při návrhu obvodů.[19] V současné době se návrh obvodů zlepšuje pomalým tempem kvůli nedostatečné organizaci známých interakcí více genů a matematických modelů. Tento problém je řešen aplikací softwaru pro počítačový design (CAD), který poskytuje multimediální reprezentaci obvodů prostřednictvím obrázků, textu a programovacího jazyka aplikovaných na biologické obvody.[20] Mezi nejznámější programy CAD patří GenoCAD, Clotho framework a j5.[21][22][23] GenoCAD používá gramatiky, které jsou buď opensource, nebo uživatelem generovaná „pravidla“, která zahrnují dostupné geny a známé genové interakce pro klonovací organismy. Clotho framework používá Standard z cihel pravidla.[20]
Reference
- ^ A b C Kobayashi, H .; Kærn, M .; Araki, M .; Chung, K .; Gardner, T. S .; Cantor, C. R .; Collins, J. J. (2004). „Programovatelné buňky: propojení přírodních a inženýrských genových sítí“. PNAS. 101 (22): 8414–8419. Bibcode:2004PNAS..101,8414 tis. doi:10.1073 / pnas.0402940101. PMC 420408. PMID 15159530.
- ^ "Syntetická biologie: FAQ". SyntheticBiology.org. Archivovány od originál dne 12. prosince 2002. Citováno 21. prosince 2011.
- ^ A b Gardner, T.s., Cantor, C.R., Collins, J. Konstrukce genetického přepínače v Escherichia coli. Příroda 403, 339-342 (20. ledna 2000).
- ^ Stanislas Leibler; Elowitz, Michael B. (leden 2000). "Syntetická oscilační síť transkripčních regulátorů". Příroda. 403 (6767): 335–338. Bibcode:2000Natur.403..335E. doi:10.1038/35002125. ISSN 1476-4687. PMID 10659856. S2CID 41632754.
- ^ Purnick, Priscilla E. M .; Weis, Ron (2009). "Druhá vlna syntetické biologie: od modulů k systémům". Nature Reviews Molecular Cell Biology. 10 (6): 410–422. doi:10.1038 / nrm2698. PMID 19461664. S2CID 200495.
- ^ International Genetically Engineered Machines (iGem) http://igem.org/Main_Page
- ^ Ro, D.-K .; Paradise, E.M .; Ouellet, M .; Fisher, K.J .; Newman, K.L .; Ndungu, J.M .; Ho, K.A .; Everyus, R.A .; Ham, T.S .; Kirby, J .; Chang, M.C.Y .; Withers, S.T .; Shiba, Y .; Sarpong, R .; Keasling, J.D. (2006). "Produkce prekurzoru antimalarického léčiva kyseliny artemisinové v připravených kvasnicích". Příroda. 440 (7086): 940–943. Bibcode:2006 Natur.440..940R. doi:10.1038 / nature04640. PMID 16612385. S2CID 3199654.
- ^ Fortman, J.L .; Chhabra, S .; Mukhopadhyay, A .; Chou, H .; Lee, T.S .; Steen, E .; Keasling, J.D. (2008). „Alternativy biopaliva k ethanolu: čerpání mikrobiální studny“. Trends Biotechnol. 26 (7): 375–381. doi:10.1016 / j.tibtech.2008.03.008. PMID 18471913.
- ^ Keasling, J.D. (2008). "Syntetická biologie pro syntetickou chemii". ACS Chem Biol. 3 (1): 64–76. doi:10.1021 / cb7002434. PMID 18205292.
- ^ Lucks, Julius B; Qi, Lei; Whitaker, Weston R; Arkin, Adam P (2008). "Směrem k rodinám škálovatelných dílů pro předvídatelný návrh biologických obvodů". Současný názor v mikrobiologii. 11 (6): 567–573. doi:10.1016 / j.mib.2008.10.002. PMID 18983935.
- ^ Elowitz, M.B .; Leibler, S. (2000). "Syntetická oscilační síť transkripčních regulátorů". Příroda. 403 (6767): 335–338. Bibcode:2000Natur.403..335E. doi:10.1038/35002125. PMID 10659856. S2CID 41632754.
- ^ Fung, E .; Wong, W.W .; Suen, J. K.; Bulter, T .; Lee, S .; Liao, J.C. (2005). "Syntetický gen - metabolický oscilátor". Příroda. 435 (7038): 118–122. Bibcode:2005 Natur.435..118F. doi:10.1038 / nature03508. PMID 15875027. S2CID 414371.
- ^ A b C Silva-Rocha, R .; de Lorenzo, V. (2008). "Těžba logických bran v prokaryotických sítích regulace transkripce". FEBS Dopisy. 582 (8): 1237–1244. doi:10.1016 / j.febslet.2008.01.060. PMID 18275855. S2CID 45553956.
- ^ A b C Buchler, N.E .; Gerland, U .; Hwa, T. (2003). „Na schématech kombinatorické transkripční logiky“. PNAS. 100 (9): 5136–5141. Bibcode:2003PNAS..100.5136B. doi:10.1073 / pnas.0930314100. PMC 404558. PMID 12702751.
- ^ Nevozhay D, Adams RM, Murphy KF, Josic K, Balázsi G (31. března 2009). „Negativní autoregulace linearizuje reakci na dávku a potlačuje heterogenitu genové exprese“. Proc. Natl. Acad. Sci. USA. 106 (13): 5123–8. Bibcode:2009PNAS..106,5123N. doi:10.1073 / pnas.0809901106. PMC 2654390. PMID 19279212.
- ^ Blake WJ, Balázsi G, Kohanski MA, Isaacs FJ, Murphy KF, Kuang Y, Cantor CR, Walt DR, Collins JJ (28. prosince 2006). „Fenotypové důsledky transkripčního šumu zprostředkovaného promotérem“. Molec. Buňka. 24 (6): 853–65. doi:10.1016 / j.molcel.2006.11.003. PMID 17189188.
- ^ Friedland, A.E .; Lu, T. K.; Wang, X .; Shi, D .; Church, G .; Collins, J.J. (2009). „Syntetické genové sítě, které se počítají“. Věda. 324 (5931): 1199–1202. Bibcode:2009Sci ... 324.1199F. doi:10.1126 / science.1172005. PMC 2690711. PMID 19478183.
- ^ Tabor, J.J .; Salis, H.M .; Simpson, Z.B .; Chevalier, A.A .; Levskaya, A .; Marcotte, E.M .; Voigt, C. A.; Ellington, A.D. (2009). „Program pro detekci syntetických hran“. Buňka. 137 (7): 1272–1281. doi:10.1016 / j.cell.2009.04.048. PMC 2775486. PMID 19563759.
- ^ Cheng, Allen A .; Lu, Timothy K. (01.01.2012). „Syntetická biologie: rozvíjející se inženýrská disciplína“. Roční přehled biomedicínského inženýrství. 14 (1): 155–178. doi:10.1146 / annurev-bioeng-071811-150118. PMID 22577777. S2CID 7319630.
- ^ A b Lux, Matthew W .; Bramlett, Brian W .; Ball, David A .; Peccoud, Jean (únor 2012). „Genetic Design Automation: Engineering Fantasy or Scientific Renewal?“. Trendy v biotechnologii. 30 (4): 120–126. doi:10.1016 / j.tibtech.2011.01.001. PMC 3073767. PMID 21310501.
- ^ „GenoCAD: CAD software pro syntetickou biologii“. www.genocad.com. Citováno 2015-10-21.
- ^ "Clotho". www.clothocad.org. Citováno 2015-10-21.
- ^ „J5“. j5.jbei.org. Citováno 2015-10-21.