ZC3H12B - ZC3H12B
| ZC3H12B | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifikátory | |||||||||||||||||||||||||
| Aliasy | ZC3H12B, CXorf32, MCPIP2, typ CCCH se zinkovým prstem obsahující 12B | ||||||||||||||||||||||||
| Externí ID | OMIM: 300889 MGI: 2442133 HomoloGene: 19395 Genové karty: ZC3H12B | ||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Ortology | |||||||||||||||||||||||||
| Druh | Člověk | Myš | |||||||||||||||||||||||
| Entrez | |||||||||||||||||||||||||
| Ensembl | |||||||||||||||||||||||||
| UniProt |
| ||||||||||||||||||||||||
| RefSeq (mRNA) | |||||||||||||||||||||||||
| RefSeq (protein) |
| ||||||||||||||||||||||||
| Místo (UCSC) | Chr X: 65,37 - 65,51 Mb | Chr X: 95,71 - 95,93 Mb | |||||||||||||||||||||||
| PubMed Vyhledávání | [3] | [4] | |||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||
| |||||||||||||||||||||||||




ZC3H12B, také známý jako CXorf32 nebo MCPIP2, je protein kódováno uživatelem gen ZC3H12B lokalizovaný na chromozomu Xq12 u lidí.
Gen
Gen ZC3H12B se skládá z 19 709 párů bází (bp) a obsahuje 5 exony. Nachází se na X chromozom na q12 na plusovém vlákně.
ZC3H12B Locus.
ZC3H12B obsahuje a ribonukleáza doména, stejně jako doména se zinkovým prstem typu CCCH. Ribonukleázy (RNasy) degradují RNA a podílejí se na procesu zrání RNA. Jsou také linií obrany proti virové RNA (D'Alessio a Riordan 1997). Zinkové prsty typu CCCH jsou spojeny s destabilizací mRNA. Ukázalo se, že zinkové prsty typu CCCH obracejí mRNA bez odstranění ocasu PolyA (Lai a Blackshear 2001). ZC3H12B a jeho paralogy Všechny ZC3H12A, ZC3H12C a ZC3H12D obsahují domény zinkových prstů typu CCCH, které byly spojeny s buněčný cyklus a přechody fází růstu u eukaryot (InterPro).
Promotér
Program Genomatix ElDorado předpovídal promotor 601 bp před genem ZC3H12B s více vazebnými místy pro transkripční faktor včetně jaderného faktoru aktivovaných T-buněk a proteinu MOK-2 spojeného se zinkovým prstem spojeným s ribonukleoproteinem ZNF239 ).
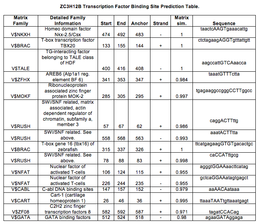
Předpovědi vazebného místa transkripčního faktoru v promotoru ZC3H12B.
mRNA
ZC3H12B obsahuje 7273 bp mRNA. Existuje pouze jeden předpokládaný přepis Aceview. Nebyly předpovídány žádné skládací vzory (Mfold). Existují introny vyříznuté ze ZC3H12B.

Single-predikovaná varianta mRNA Aceview v ZC3H12B.

Protein
ZC3H12B je pravděpodobná ribonukleáza obsahující domény zinkových prstů typu CCCH a domény ribonukleázy. Protein 836 aminokyselin má předpokládanou molekulovou hmotnost 94,2 kdal. Neobsahuje signální peptid ani transmembránovou oblast. PSORTII předpovídal 65,2% pravděpodobnost jaderného umístění. Zinkové prsty a ribonukleázy typu CCCH jsou pravděpodobně umístěny v jádru pro štěpení RNA a konkrétně štěpení RNA vlásenkami (Boysen a Hearn 2008).
Strukturální charakteristiky
The sekundární struktura bílkovin je směsí alfa helixy a beta řetězce. Dvě dosud identifikované domény jsou domény zinkových prstů ribonukleázy a CCCH typu.
Níže je zobrazena konzervovaná doména paralogu ZC3H12B, Mcpip1 (nebo ZC3H12A). Ve srovnání struktury BLAST byla 82% shoda identity s 24% pokrytím dotazu, s předpokládanou e-hodnotou 2e-118,82% shoda identity je dostatečná k provedení srovnání domény konzervované zinkovým prstem ZC3H12B a Mcpip1 (ZC3H12A), která se předpokládá, že se budou skládat z beta řetězců a alfa šroubovic.

Cn3D vykreslování konzervované domény Mcpip1 s motivem zinkového prstu.

Posttranslační modifikace
Program Phobius předpovídal umístění necytoplazmatického proteinu. NetPhos 2.0. předpokládalo 63 fosforylačních míst v ZC3H12B, která jsou označena na koncepční translaci. 1.2. předpověděl tři místa připojení 0-Beta-GlcNAc, která soutěží s místy fosforylace. 0-Beta-GlcNAc je pravděpodobně jediný typ glykosylace vyskytující se v jádru a / nebo cytoplazmě buněk. Existuje pozoruhodná souvislost mezi aktivací antigenu lymfocyty a dynamickou 0-B-glykosylací v jaderných proteinech (Hart a Akimoto). NetNGlyc předpovídal glykosylační místa; tato místa však byla vyloučena, protože protein je pravděpodobně jaderný a nepodstoupil by tuto formu glykosylace. Na N-konci proteinu nebyla žádná předpovězená acetylační místa. To je neobvyklé, protože přibližně 85% lidských proteinů je acetylováno na N-konci pro syntézu, stabilizaci a lokalizaci proteinů (Van Damm et al.). Nejsou přítomny žádné shluky kladných, záporných nebo smíšených nábojů. Nebyly zjištěny žádné hydrofobní segmenty (SAPS SDSC Biology Workbench). MitoProtII nezjistil žádné signály exportu mitochondrií. Tyto posttranslační testy naznačují, že protein je umístěn v jádru a prochází dynamickou fosforylací a modifikacemi 0-Beta-GlcNAc.
Vývoj
Vybrané domény ZC3H12B jsou konzervovány u většiny obratlovců, členovců a kroužkovců. V doménách bakterie nebo archea nejsou konzervované domény. U kvasinek, rostlin nebo protistů nebyly významně konzervované domény.
Paralogy
Tam jsou tři paralogy ZC3H12B, které jsou ve stejné rodině zinkových prstů typu CCCH, všechny, které si na základě analýzy BLAST (NCBI) udržují více než 50% identitu se ZC3H12B.
| název | Druh | NCBI přístupové číslo | Délka (AA) | Proteinová identita |
|---|---|---|---|---|
| ZC3H12B | Homo sapiens | NM_001010888.3 | 836aa | 100% |
| ZC3H12A | Homo sapiens | NM_025079.2 | 599aa | 68% |
| ZC3H12C | Homo sapiens | NM_033390.1 | 883aa | 53% |
| ZC3H12D | Homo sapiens | NM_207360.2 | 527aa | 61% |
Ortology
ZC3H12B je konzervován u savců, ptáků, hmyzu a hlístic (BLAST). V tabulce níže je uveden souhrn ortology ZC3H12B u lidí.
| Druh | Obecný název druhu | Divergence (MYA) | NCBI přístupové číslo (protein) | Délka (aminokyseliny) | Proteinová identita | Podobnost | |
|---|---|---|---|---|---|---|---|
| Homo sapiens | Člověk | n / a | NP_001010888.3 | 836aa | 100% | 100% | |
| Pan paniscus | Šimpanz | 6.3 | XP_003816967.1 | 836aa | 99% | 99% | |
| Pongo abelii | Orangutan | 15.7 | XP_002831786.1 | 836aa | 99% | 99% | |
| Macaca mulatta | Rhesus opice | 29 | XP_002806307.1 | 836aa | 99% | 99% | |
| Callithrix jacchus | Kočkodan | 42.6 | XP_002762992.2 | 836aa | 98% | 98% | |
| Mus musculus | Myš | 92.3 | NP_001030079.2 | 835aa | 91% | 94% | |
| Sus scrofa | Prase | 94.2 | XP_003360389.1 | 836aa | 93% | 96% | |
| Gallus gallus | Kuře | 296 | XP_003641177.1 | 837aa | 77% | 85% | |
| Chrysemys picta bellii | Malovaná želva | 296 | XP_005279572.1 | 838aa | 78% | 86% | |
| Oryzias latipes | Medaka | 400.1 | XP_004076599.1 | 845aa | 67% | 77% | |
| Gadus morhua | Treska obecná | 400.1 | AFK76491.1 | 842aa | 29% | 44% | |
| Danio rerio | Zebrafish | 400.1 | XP_001342172.3 | 982aa | 68% | 77% | |
| Petromyzon marinus | Mihule | 535.7 | ABO21295.1 | 222aa | 44% | 58% | |
| Branchiostoma floridae | Lancelet | 713.2 | XP_002598834.1 | 492aa | 66% | 79% | |
| Ciona intestinalis | Váza tunikát | 722.5 | XP_002125834.1 | 863aa | 54% | 66% | |
| Strongylocentrotus purpuratus | Fialový mořský ježek | 742.9 | XP_787030.3 | 974aa | 58% | 72% | |
| Aplysiomorpha californica | Mořský zajíc | 782.7 | XP_005113312.1 | 1269aa | 51% | 69% | |
| Drosophila grimshawi | Havajská ovocná muška | 782.7 | XP_001994140.1 | 548aa | 51% | 69% | |
| Anopheles gambiae | Komár | 782.7 | XP_321880 | 637aa | 59% | 75% | |
| Apis mellifera | Včelí med | 782.7 | XP_397264 | 652aa | 58% | 73% | |
| Caenorhabditis elegans | Kulatý červ (hlístice) | 937.5 | NP_491985.4 | 634aa | 46% | 64% |
Výraz a funkce
Microarrays v normální tkáňové expresi profilování ukázalo zvýšenou expresi genu ve slinivce břišní, prostatě, mozku, míchě a brzlíku (GEO). Na rozdíl od jeho paralogy, není exprimován v tkáních aktivovaných makrofágy, což naznačuje paralogní vztah k zánětlivé odpovědi (Liang et al. 2008). ZC3H12B je přechodně exprimován v tkáních mozku, brzlíku a varlat (EST).
Interakce
Předpokládané interakce společnosti Ingenuity Systems neprokázaly žádné molekuly zaměřené na léky v cestě a žádné známé cíle léčiv. Uvedené hlavní funkce a nemoci byly rakovina, poškození organismu a abnormality, onemocnění reprodukčního systému. Několik miRNA byly předpovídány interakce. Předpovězené cíle miRNA je ještě třeba porovnat se sekvencí ZC3H12B a není jasné, zda mezi nimi existuje interakce. K testování nových interakcí protein / chromatin se ZC3H12B lze v budoucnu použít testy, jako je Forster Resonance Energy Transfer (FRET), koimunooprecipitace, dvouhybridní screening, hydropatická komplementarita, cluster-microarray a ChiP.
Klinický význam
Odstranění lokusu Xq12 mělo za následek několik poruch, jako je necitlivost na androgeny, náchylnost k rakovina prostaty, spinální a bulbární svalová atrofie Kennedyho amentální retardace; mezi těmito chorobami a ZC3H12B (NCBI) však nebyla nalezena žádná souvislost.
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000102053 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000035045 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
Další čtení
- Liang J, Wang J, Azfer A, Song W, Tromp G, Kolattukudy PE, Fu M (březen 2008). „Nová rodina proteinů s CCCH-zinkovými prsty reguluje prozánětlivou aktivaci makrofágů“. The Journal of Biological Chemistry. 283 (10): 6337–46. doi:10.1074 / jbc.m707861200. PMID 18178554.
- Van Damme P, Hole K, Pimenta-Marques A, Helsens K, Vandekerckhove J, Martinho RG, Gevaert K, Arnesen T (červenec 2011). „NatF přispívá k evolučnímu posunu N-terminální acetylace proteinu a je důležitý pro normální segregaci chromozomů“. PLoS Genetics. 7 (7): e1002169. doi:10.1371 / journal.pgen.1002169. PMC 3131286. PMID 21750686.