Thalassiosira - Thalassiosira
| Thalassiosira | |
|---|---|
 | |
| Fosilní druhy Thalassiosira | |
| Vědecká klasifikace | |
| Doména: | |
| (bez hodnocení): | |
| Nadkmen: | |
| Třída: | |
| Objednat: | |
| Rodina: | |
| Rod: | Thalassiosira P.T. Cleve, 1873 emend. Hasle, 1973 |
| Druh | |
Viz text | |
Thalassiosira je rod centrických rozsivek, který zahrnuje více než 100 mořských a sladkovodních druhů. Jedná se o různorodou skupinu fotosyntetických eukaryot, které tvoří důležitou součást mořských a sladkovodních ekosystémů, v nichž jsou klíčovými primárními producenty a jsou nezbytné pro cyklování uhlíkem. [1]
Thalassiosira je rozmanitý rod, nicméně jeden druh v rodu, T. pseudonanazískal zvláštní význam jako první mořský fytoplankton, u kterého byl sekvenován genom. T. pseudonana se od té doby stal klíčovým modelovým organismem pro studium fyziologie rozsivek. The T. pseudonana genom odhalil nové geny pro intracelulární obchodování a metabolismus v rozsivkách [2]. Tento druh byl znovu použit k vývoji metod pro genetickou manipulaci rozsivek [3] a pro studium biomineralizace oxidu křemičitého [4].
Pozadí
Thalassiosira byl poprvé popsán v roce 1873 P.T. Cleve [5]. Rod byl poté charakterizován transmisní elektronovou mikroskopií v padesátých letech a skenovací elektronovou mikroskopií v šedesátých letech, což vedlo k lepšímu pochopení definujících morfologických rysů rodu a následnému rozpoznání více než 100 druhů [6]
Druh Thalassiosira, T. pseudonana, byl vybrán jako první mořský eukaryotický fytoplankton, který podstoupil celé genomové sekvenování, kvůli jeho malému genomu 34 MB. Jako výsledek, T. pseudonana sloužil jako modelový organismus pro porozumění biologii rozsivek. Sekvenování celého genomu, transkriptomika a proteomika genomu T. pseudonana odhalily nové cesty pro biogenezi křemíku, stresovou reakci fosforu, intracelulární transport a metabolismus v mořských rozsivkách [2] [7][8]. Nedávné genetické studie využívající genové sekvence ribozomální RNA také dokázaly lépe rozlišit a klasifikovat druhy Thalassiosira [1].
Popis
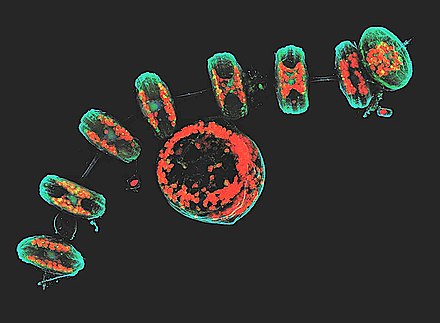
Tento konfokální obraz ukazuje rozsivky buněčná stěna (tyrkysová), chloroplasty (Červené), DNA (modrý), membrány a organely (zelený).
Thalassiosira přicházejí v různých tvarech, od krabicových až po válcové, diskoidní nebo sférické [9]. Nějaký Thalassiosira buňky se nacházejí samostatně, zatímco jiné tvoří řetězce. Thalassiosira mají buněčnou stěnu vyrobenou z oxidu křemičitého, známého jako frustule [5]. Thalasiosira skrývají několik diskoidních plastidů a kruhový ventil, který obsahuje póry uspořádané v řadách nebo obloucích, které se otevírají ven. Okraj pláště ventilu je lemován řadou pásů. Různé druhy Thalassiosira lze identifikovat podle morfologických charakteristik jejich areol a procesů na chlopni [10]. Během tvorby kolonií Thalassiosira uvolňuje chitinová vlákna strutted procesy známými jako fultoportulae. Vytlačováním chitinových vláken a tím zvyšováním odporu může Thalassiosira zpomalit rychlost jejich klesání [2].
Stanoviště a ekologie
Thalassiosira zabírají různá stanoviště, mořská i sladkovodní. Za zmínku stojí, že jsou životně důležitými primárními producenty v mírných a polárních oblastech [1]. Thalassiosira mohou prospívat při nízké teplotě a světle, stejně jako ve smíšených vodách, a proto jsou na jaře velkou částí květů rozsivky v mírných oblastech, jako jsou kanadské a aljašské vody [10]. Druhy tohoto rodu jsou také schopné sestavovat obranná vlákna proti zooplanktonu, což jim umožňuje přežít predaci, která za normálních okolností udržuje květy fytoplanktonu pod kontrolou.
Thalassiosira Druhy jsou rozmanité jak ve své ekologii, tak ve fyziologii, s různými mechanismy pro ukládání dusíku nebo požadavky na železo. Koncentrace železa, teplota a dostupnost makroživin byly identifikovány jako důležité faktory pro složení Thalassiosira společenstva druhů v mořských vodách [1].
Genetika
Fylogenetické studie založené na 18S rRNA genech odhalily několik subtypů uvnitř Thalassiosira, celková parafyletická skupina v rodině Thalassiosirales [11]. Vztahy druhů uvnitř subtypů je třeba ještě objasnit.
Posloupnost T. pseudonana genom odhalil řadu funkcí, které odlišují rozsivky od jiných eukaryot. Například transkripční faktory tepelného šoku tvoří většinu transkripčních faktorů v T. pseudonana genom, i když u jiných eukaryot jsou méně časté [2]. Také se odlišuje od ostatních eukaryotických genomů relativní absencí receptorových kináz a receptorů spojených s G proteinem. Specifičtější pro biologii rozsivek bylo pochopení biochemie křemíku u rozsivek vylepšeno objevem genů podílejících se na vychytávání kyseliny křemičité a proteinů zapojených do vezikul pro srážení oxidu křemičitého.
Překvapivé zjištění z T. pseudonana genomem byla přítomnost genů kódujících enzymy pro kompletní cyklus močoviny, což bylo u fotosyntetického eukaryotu bezprecedentní [2]. Je známo, že se používá dvojiatomů, které využívají enzym ureázy ke katalyzování rozpadu močoviny, a proto se neočekává, že budou potřebovat mechanismy, které by jej vylučovaly jako odpad. Kupodivu T. pseudonana močovinový cyklus se napájí do dalších metabolických drah, které přispívají k biosyntéze bílkovin a případně k ukládání energie.
Životní cyklus
Thalassiosira může podstoupit asexuální a sexuální reprodukci v procesech sdílených jinými rozsivkami [12]. Během nepohlavní reprodukce se nadřazená buňka dělí na dvě dceřiné buňky nerovné velikosti - jednu o velikosti stejné jako mateřská a jednu menší. Toto omezení velikosti během mitotického dělení je způsobeno přítomností tuhé buněčné stěny oxidu křemičitého. Výsledkem je, že při více buněčných děleních se zmenší velikost buněk každé dceřiné buňky. Abychom zvládli zmenšování velikosti buněk, Thalassiosira může přejít na sexuální reprodukci, která je vyvolána řadou faktorů prostředí, které nejsou dobře pochopeny, jakmile buňky dosáhnou kriticky malé velikosti [13]. Při pohlavním rozmnožování se spermie a vejce, které mohou vzniknout ze stejné buňky, spojí a vytvoří diploidní zygotu, která se označuje jako auxospora. Toto potomstvo pak může vyjít z rodičovské frustule (křemičité stěny) a rekonstruovat svou vlastní buněčnou stěnu, čímž se stane buňkou větší velikosti.
Fosilní historie
Geologický záznam Thalassiosira pochází z doby před 13,82 miliony let [9]. Fosilní záznam o sladkovodních i mořských Thalassiosira organismy je značný, ačkoli sladkovodní sběr byl charakterizován extenzivněji [14].
Praktický význam
T. pseudonana je zvláště užitečný pro molekulární studie kvůli své malé velikosti genomu. Odhalila nové cesty pro biosyntézu oxidu křemičitého, které zahrnují absorpci kyseliny monosilicic z mořského prostředí, intracelulární transport do specializovaného vezikuly a speciální enzymy a peptidy pohánějící syntézu oxidu křemičitého [4].
Biosyntéza oxidu křemičitého, pro kterou druh T. pseudonana přišel sloužit jako modelový organismus pro studium, byl zvláště zajímavý v kontextu nanotechnologie strojírenského křemene [2]. Například upravený T. pseudonana byl prozkoumán jako vezikul pro dodávání léků v léčbě rakoviny [15].
Thalassiosira, kromě jiných rozsivek, byly také považovány za potenciálně užitečné zdroje lipidů pro biopaliva. Konkrétně T. weissflogii Bylo prokázáno, že snižuje svoji syntézu oxidu křemičitého a současně reguluje produkci triacylglycerolů při kultivaci v podmínkách omezujících dusík. [16].
Vědecká klasifikace
Chromista, Harosa, Heterokonta, Ochrophyta, Bacillariophyceae, Coscinodiscophycidae, Thalassiosiranae, Thalassiosirales, Thalassiosiraceae, Thalassiosira [17]
|
Reference
- ^ A b C d Dreux Chappell, P., Whitney, L. A. P., Haddock, T. L., Menden-Deuer, S., Roy, E. G., Wells, M. L. a Jenkins, B. D. (2013). Thalassiosira spp. složení komunity se mění v reakci na chemické a fyzikální působení na severovýchodě Tichého oceánu. Frontiers in Microbiology, 4 (SEP), 273. https://doi.org/10.3389/fmicb.2013.00273
- ^ A b C d E F Armbrust, E. V. (2004). Genom Diatom Thalassiosira pseudonana: Ekologie, evoluce a metabolismus. Science, 306 (5693), 79–86. https://doi.org/10.1126/science.1101156
- ^ Poulsen, N., Chesley, P. M. a Kröger, N. (2006). Molekulárně genetická manipulace rozsivky Thalassiosira pseudonana (Bacillariophyceae). Journal of Phycology, 42 (5), 1059–1065. https://doi.org/10.1111/j.1529-8817.2006.00269.x
- ^ A b Sumper, M., & Brunner, E. (2008, 23. května). Biomineralizace oxidu křemičitého v rozsivkách: Modelový organismus Thalassiosira pseudonana. ChemBioChem. https://doi.org/10.1002/cbic.200700764
- ^ A b Miranda, S. V., Guiry, M. D. a Guiry, G. (2015, 19. června). Thalassiosira Cleve, 1873: Algaebase. Citováno 30. března 2020 z https://www.algaebase.org/search/genus/detail/?genus_id=43768&-session=abv4:AC1F24C807fd0012FBNw3B92519A
- ^ (Bacillariophyta) druhy zřídka zaznamenané v brazilských pobřežních vodách. Braz. J. Biol (svazek 69).
- ^ Dyhrman, S. T., Jenkins, B. D., Rynearson, T. A., Saito, M. A., Mercier, M. L., Alexander, H.,… Heithoff, A. (2012). Transkriptom a proteom rozsivky thalassiosira pseudonana odhalují rozmanitou stresovou reakci fosforu. PLoS ONE, 7 (3), e33768. https://doi.org/10.1371/journal.pone.0033768
- ^ Mock, T., Samanta, M. P., Iverson, V., Berthiaume, C., Robison, M., Holtermann, K.,… Armbrust, E. V. (2008). Profilování exprese celého genomu mořské rozsivky Thalassiosira pseudonana identifikuje geny zapojené do křemíkových bioprocesů. Sborník Národní akademie věd Spojených států amerických, 105 (5), 1579–1584. https://doi.org/10.1073/pnas.0707946105
- ^ A b Diatom - Thalassiosira. (n.d.). Citováno 27. února 2020, z https://algaeresearchsupply.com/pages/the-diatom-thalassiosira
- ^ A b Harris, A., Medlin, L., Lewis, J., Jones, K., D Harris, A. S., Medlin, L. K., & Jones, K. J. (1995). Druh Thalassiosira (Bacillariophyceae) ze skotského mořského jezera. European Journal of Phycology, 30 (2), 117–148. https://doi.org/10.1080/09670269500650881
- ^ Hoppenrath, M., Beszteri, B., Drebes, G., Halliger, H., Van Beusekom, J. E. E., Janisch, S., & Wiltshire, K. H. (2007). Druhy Thalassiosira (Bacillariophyceae, Thalassiosirales) v Severním moři v Helgolandu (německé zátoce) a Sylt (severofríské Waddenské moře) - první přístup k hodnocení rozmanitosti. European Journal of Phycology, 42 (3), 271–288. https://doi.org/10.1080/09670260701352288
- ^ Moore, E. R., Bullington, B. S., Weisberg, A. J., Jiang, Y., Chang, J., & Halsey, K. H. (2017). Morfologické a transkriptomické důkazy indukce sexuální reprodukce reprodukcí amoniakem u Thalassiosira pseudonana a dalších centrických rozsivek. PLOS ONE, 12 (7), e0181098. https://doi.org/10.1371/journal.pone.0181098
- ^ Armbrust, E. V. (1999). Identifikace nové genové rodiny vyjádřené během nástupu sexuální reprodukce v centrickém rozsivce Thalassiosira weissflogii. Aplikovaná a environmentální mikrobiologie, 65 (7), 3121–3128. https://doi.org/10.1128/aem.65.7.3121-3128.1999
- ^ Alverson, A. J. (2014). Načasování mořských – sladkovodních přechodů v rozsivce řádu Thalassiosirales. Paleobiologie, 40 (1), 91–101. https://doi.org/10.1666/12055
- ^ Delalat, B., Sheppard, V. C., Rasi Ghaemi, S., Rao, S., Prestidge, C. A., McPhee, G.,… Voelcker, N. H. (2015). Cílené podávání léků pomocí geneticky upravené křemeliny. Nature Communications, 6 (1), 1–11. https://doi.org/10.1038/ncomms9791
- ^ D’Ippolito, G., Sardo, A., Paris, D., Vella, F. M., Adelfi, M. G., Botte, P., ... Fontana, A. (2015). Potenciál metabolismu lipidů v mořských rozsivkách pro výrobu biopaliv. Biotechnology for Biopalels, 8 (1), 28. https://doi.org/10.1186/s13068-015-0212-4
- ^ Thalassiosira pseudonana Hasle & Heimdal, 1970. (2005, 30. března). Citováno 31. března 2020, z http://www.marinespecies.org/aphia.php?p=taxdetails&id=148934
Další reference
- Růst Thalassiosira fluviatilis omezený na živiny a světlo v kontinuální kultuře s důsledky pro růst fytoplanktonu v oceánu. EA Laws, TT Bannister - Limnology and Oceanography, 1980
- Vliv chemické chemie železa na příjem železa pobřežní rozsivkou Thalassiosira weissflogii. MA Anderson, FMM Morel, Limnology and Oceanography, 1982
externí odkazy
 Údaje týkající se Thalassiosira na Wikispecies
Údaje týkající se Thalassiosira na Wikispecies Média související s Thalassiosira na Wikimedia Commons
Média související s Thalassiosira na Wikimedia Commons
| Tento rozsivka článek je a pahýl. Wikipedii můžete pomoci pomocí rozšiřovat to. |