Photobacterium profundum - Photobacterium profundum
| Photobacterium profundum | |
|---|---|
 | |
| Konfokální obrázek P. profundum Strain SS9 exprimující a zelený fluorescenční protein. Bar: 10 μm | |
| Vědecká klasifikace | |
| Království: | |
| Kmen: | |
| Třída: | |
| Objednat: | Vibrionales |
| Rodina: | |
| Rod: | |
| Druh: | P. profundum |
| Binomické jméno | |
| Photobacterium profundum | |
Photobacterium profundum je hluboké moře Gammaproteobacterium, patřící do rodiny Vibrionaceae a rod Photobacterium. Stejně jako ostatní členové tohoto rodu, P. profundum je mořský organismus a má dva kruhy chromozomy.[1] P. profundum je gramnegativní prut se schopností růstu při teplotách od 0 ° C do 25 ° C a tlacích od 0,1 MPa do 70 MPa v závislosti na kmeni. Má požadavek na sůl, je schopen metabolizovat širokou škálu jednoduchých a komplexních sacharidy a má dva bičíky systémy. Buňky mají tyčový tvar, délku 2–4 μm a šířku 0,8–1,0 μm, s jedním nezakrytým bičíkem.[2] Tato bakterie byla původně izolována v roce 1986 z Suluské moře a v současné době existují 4 kultivované kmeny divokého typu P. profundum, (kmeny SS9, 3TCK, DJS4 a 1230).[3]
P. profundum kmen SS9 má optimální růst při 15 ° C a 28 MPa, takže je a psychrofil a a piezofil. P. profundum kmen 3TCK, izoláty z San Diego Bay,[4] optimálně roste při 9 ° C a 0,1 MPa a P. profundum kmen DSJ4, izolovaný z Ryukyu příkop vypnutý z Japonsko v hloubce 510 m optimálně roste při 10 ° C a 10 MPa.[2] Na základě 16S rRNA sekvence P. profundum úzce souvisí s rodem Vibrio, nejvýznamnějším druhem je lidský patogen Vibrio cholerae.
U kmene SS9 se ukázalo, že několik genů stresové odezvy je regulováno v reakci na atmosférický tlak, mezi ně patří htpG, dnaK, dnaJ a groEL.[1] Druhy a množství řetězců mastných kyselin v buněčné membráně také reagují na změny tlaku a teploty.[5] Při nízké teplotě a vysokém tlaku napětí SS9 zvyšuje hojnost mono- a polynenasycené mastné kyseliny. To má za následek zvýšení tekutosti membrány snížením náplně řetězců mastných kyselin, což má za následek spíše strukturu tekutých krystalů v membráně než gelovou strukturu.[5] The protein vnější membrány Ukázalo se, že OmpH je regulován nahoru při zvýšených tlacích, opak je pravdou pro protein OmpL vnější membrány, který je regulován nahoru v reakci na nízké tlaky.[6]
V roce 2005 Vezzi et al. zveřejnil sekvence genomu pro P. profundum kmen SS9. Genom P. profundum skládá se z kruhového chromozomu 4,1 Mbp, malého kruhového chromozomu 2,2 Mbp a kruhového 80 kbp plazmid. Kmen SS9 má 14 genů ribozomální RNA (rRNA) na chromozomu 1 a 1 na chromozomu 2; toto je největší počet genů rRNA nalezených v jakékoli bakterii. Chromozom 1 se skládá převážně z genů, které jsou nezbytné pro růst, zatímco chromozom 2 se jeví jako velký plazmid, který v evolučním časovém měřítku získal několik transponovatelné prvky.[4] V genomu P. profundum existuje velké množství otevřené čtecí rámce (ORF), které jsou jedinečné pro SS9 a nebyly nalezeny u jiných členů rodiny Vibrionaceae.
Sekvence genomu také zvýraznila celou Stickland cesta pro fermentaci aminokyseliny; toto bylo poprvé, co byla tato cesta identifikována v aerobní bakterie. V této bakterii jsou také přítomny dvě kompletní F1F0 ATP syntázové dráhy (jedna na každém chromozomu): to může vysvětlovat její schopnost produkovat ATP při vysokém i nízkém tlaku.
Na tuto práci navázal v roce 2005 další dokument Campanaro et al. který podrobně microarray práce srovnávající genovou expresi při suboptimálních, optimálních a supraoptimálních teplotách a tlaku pro kmeny SS9, 3TCK a DSJ4.[4] Campanaro a kol. ukázalo, že v genomu 3TCK existuje 544 ORF divergentních nebo chybějících a 562 ORF divergentních nebo chybějících v chromozomech DSJ4 ve srovnání s chromozomy SS9. Tento dokument také zdůraznil, že 3TCK postrádá systém laterálních bičíků, který je v SS9 regulován při zvýšeném tlaku, stejně jako absence 3 fág související regiony z 3TCK a 4 regiony související s fágem z DSJ4.[4][7]
Transkripční krajina divokého typu kmene DB110 a toxR mutantní TW30 byly zkoumány pomocí sekvenování nové generace.[8] ToxR je transmembrána Protein vázající DNA poprvé objeven v Vibrio cholerae, kde reguluje značný počet genů zapojených do adaptace na životní prostředí a virulence. v P. profundum hojnost a aktivita tohoto proteinu je ovlivněna hydrostatickým tlakem a jeho role souvisí s regulací genů tlakově závislým způsobem.[9] Výsledky získané z RNA sekvence experimenty odhalily komplexní vzor exprese se skupinou 22 genů, které mají profily exprese podobné OmpH, což je protein vnější membrány transkribovaný v reakci na vysoký hydrostatický tlak.[10] Kromě toho RNA-seq umožnila hlubokou charakterizaci transkripční krajiny, která vedla k identifikaci 460 domnělých malá RNA genů a detekce 298 genů kódujících proteiny dříve neznámých. Genomová předpověď pro operon Struktura, stránky pro zahájení a ukončení transkripce, odhalily neočekávaně vysoký počet genů (992) s velkými 5’-UTR, dostatečně dlouhými k uložení cis-regulační RNA struktury, což naznačuje korelaci mezi velikostí intergenní oblasti a délkou UTR.
CSLM obrázek počátečních fází biofilm tvorba v Photobacterium profundum SS9 za použití živých / mrtvých skvrn. Tlak = 28 MPa Teplota = 15 stupňů C Bar = 10 mikrometrů

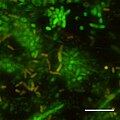
CSLM Photobacterium profundum SS9 exprimující GFP. Bar = 10 mikrometrů


Reference
- ^ A b Vezzi, A. a kol., Life at depth: Photobacterium profundum genomová sekvence a analýza exprese. Science, 2005. 307 (5714): str. 1459-61.
- ^ A b Nogi, Y., N. Masui a C. Kato, Photobacterium profundum sp. nov., nový, mírně barofilní bakteriální druh izolovaný z hlubinného sedimentu. Extremophiles, 1998. 2 (1): str. 1-7.
- ^ DeLong, E.F., Adaptace hlubinné bakterie na hlubinné prostředí 1986.
- ^ A b C d Campanaro, S, Vezzi, A, Vitulo, N, Lauro, FM, D'Angelo, M, Simonato, F, Cestaro, A, Malacrida, G, Bertoloni, G, Valle, G, Bartlett, DH., Laterally přenesené prvky a adaptace na vysoký tlak u kmenů Photobacterium profundum. BMC Genomics, 2005. 6: str. 122.
- ^ A b Allen, E.E., D. Facciotti a D.H. Bartlett, Mononenasycené, ale ne polynenasycené mastné kyseliny, jsou potřebné pro růst hlubinné bakterie Photobacterium profundum SS9 za vysokého tlaku a nízké teploty. Appl Environ Microbiol, 1999. 65 (4): str. 1710-20.
- ^ Bartlett, D.H. a T.J. Exprese genu Welch, ompH je regulována mnoha podněty prostředí kromě vysokého tlaku v hlubinné bakterii Photobacterium species kmen SS9. J Bacteriol, 1995. 177 (4): str. 1008-16.
- ^ Campanaro, S, Vezzi, A, Vitulo, N, Lauro, FM, D'Angelo, M, Simonato, F, Cestaro, A, Malacrida, G, Bertoloni, G, Valle, G, Bartlett, DH., Laterally přenesené prvky a adaptace na vysoký tlak u kmenů Photobacterium profundum. BMC Genomics, 2005. 6: str. 122.
- ^ Campanaro, S, DePascale, F, Telatin A, Schiavon R, Bartlett, DH, Valle, G. Transkripční krajina hlubinné bakterie Photobacterium profundum jak u mutantu toxR, tak u jeho rodičovského kmene. BMC Genomics, 2012 13: str. 567. doi: 10,1186 / 1471-2164-13-567.
- ^ DiRita, VJ, Mekalanos, JJ. Periplazmatická interakce mezi dvěma membránovými regulačními proteiny, ToxR a ToxS, vede k signální transdukci a transkripční aktivaci. Cell, 1991. 64 (1): str. 29–37.
- ^ Welch, TJ, Bartlett, DH, Identifikace regulačního proteinu požadovaného pro expresi genu reagujícího na tlak v hlubinné bakterii Photobacterium druhový kmen SS9. Mol Microbiol, 1998. 27 (5): str. 977–985.