Niche (proteinový strukturní motiv) - Niche (protein structural motif)
V oblasti protein strukturální motivy, výklenky jsou tři nebo čtyři aminokyselina zbytkové vlastnosti, ve kterých jsou CO skupiny hlavního řetězce přemostěny kladně nabitými nebo δ + skupinami.[1][2][3] Skupiny δ + zahrnují skupiny se dvěma vodíková vazba donorové atomy, jako je NH2 skupiny a molekuly vody. V typických proteinech 7% aminokyselinových zbytků patří do výklenků vázaných na skupinu 5 +, zatímco dalších 7% má konformaci, ale žádný kationtový je detekována přemosťovací skupina.
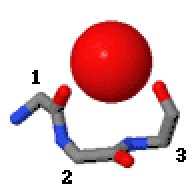
Niky jsou dvou druhů, které se rozlišují jako výklenek3 (3 zbytky, i až i + 2) a výklenek4 (4 zbytky, i až i + 3). V motivu výklenku 3 jsou karbonylové skupiny vázající 5 + ze zbytků i a i + 2, zatímco v motivu výklenku 4 jsou ze zbytků i a i + 3.
Nika3 má α konformaci pro zbytek i + 1 a β konformaci pro zbytek i + 2; niche4 má α konformaci pro zbytky i + 1 a i + 2 a β konformaci pro zbytek i + 3.
Nika se běžně vyskytuje u C-konec z α-šroubovice, a zejména z 310 šroubovice.
Kovové ionty, které se vyskytují navázané na mezery v proteinech, jsou Na +, K +, Ca ++ a Mg ++. Proteiny s regulačními kationty často používají mezery pro vázání kovů (trombin, Na +; anexin, Ca ++; pyruvátdehydrogenáza, K +).
Hlavním transportérem kationtů v buňkách je vápenatou ATPázu.[4] V krystalových strukturách vázaných na Ca ++ jsou dva ionty vápníku vedle sebe uvnitř transmembránová doména jsou považovány za v polovině transportní fáze. Stejně jako být vázán různými postranními řetězci karbonylové skupiny, jeden z těchto iontů vápníku je vázán nika3 / nika4 (obě v jednom motivu) na zbytcích 304–307 na C-konci α-šroubovice.
A lysin postranní řetězec v nukleárním exportním receptoru CRM1 je specificky uznán specifickou konformací, kterou je třeba přijmout jako klíčovou součást jaderný exportní signál proteinů opouštějících jádro.[5]
Ion sodíku ve fluoridovém kanálu Fluc se nachází na ose dyadu dimeru, který je čtyřstěnně vázán dvěma výklenky4, jednou z každé podjednotky.[6]
Oblast linkeru Hsp70 interdomény 10 zbytků umožňuje alosterickou komunikaci mezi dvěma složenými doménami. N-koncová část linkeru má strukturu výklenku4, která je vázána na vodu.[7]
V toxinu Scorpion BeM9 se postranní řetězec argininu 60 váže na karbonyly zbytků 61 a 63 jako výklenek3. Motiv, jehož ztráta mění specificitu proteinu pro napěťově řízené sodíkové kanály, se nazývá „ruka argininu“[8]. Trochu neobvyklé úhly vzepětí pro niku3 jsou proto, že tato nika3 pojme dvě oddělené NH skupiny od skupiny guanidino argininu.
Další malý tripeptidový motiv, který váže kationty nebo skupiny 5 + prostřednictvím skupin CO v hlavním řetězci, se nazývá Catgrip.
Reference
- ^ Torrance, GM; Leader DP (2009). „Nový hlavní motiv řetězce v proteinech přemostěných kationickými skupinami: Niche“. Journal of Molecular Biology. 385 (4): 1076–1086. doi:10.1016 / j.jmb.2008.11.007. PMID 19038265.
- ^ Regad, L; Martin J (2011). „Disekční proteinové smyčky se statistickým skalpelem naznačují funkční implikaci některých strukturních motivů“. BMC bioinformatika. 12 (1): 247. doi:10.1186/1471-2105-12-247. PMC 3158783. PMID 21689388.
- ^ Cianci, M; Tomaszewski (2010). "Krystalografická analýza účinků opačného působení na enzymatické působení subtilisinu v acetonitrilu". Journal of the American Chemical Society. 132 (7): 2293–2300. doi:10.1021 / ja908703c. PMID 20099851.
- ^ Toyoshima, C; Mizutani (2004). "Krystalová struktura vápníkové pumpy s navázaným analogem ATP". Příroda. 430 (6999): 529–535. doi:10.1038 / nature02680. PMID 15229613.
- ^ Fung, HYJ; Fu S-C; Chook YM (2017). „Jaderný exportní receptor CRM1 rozpoznává různé konformace v jaderných exportních signálech“. eLife. 6: e23961. doi:10,7554 / eLife.23961. PMC 5358978. PMID 28282025.
- ^ Stockbridge, RB; Kolmakova-Partensky L; Shane T (2015). „Krystalové struktury dvousložkového fluoridového iontového kanálu“. Příroda. 525: 548–551. doi:10.1038 / příroda14981. PMC 4876929.
- ^ Angličtina, CA; Sherman W; Meng W (2017). „Propojovací doména Hsp70 je dynamický přepínač, který umožňuje alosterickou komunikaci mezi dvěma strukturovanými doménami“. J Biol Chem. 292: 14765–14774. doi:10.1074 / jbc.M117.789313. PMC 5592658. PMID 28754691.
- ^ Kuldyushev, NA; Mineev KS; Berkut AA (2018). "Rafinovaná struktura BeM9 odhaluje arginovou ruku, motiv toxinů štíra ovlivňující sodíkové kanály". Proteiny. 86: 1117–1122. doi:10,1002 / prot.25583.
externí odkazy
- ^ Leader, DP; Milner-White (2009). „Motivované proteiny: Webová aplikace pro studium malých trojrozměrných proteinových motivů“. BMC bioinformatika. 10 (1): 60. doi:10.1186/1471-2105-10-60. PMC 2651126. PMID 19210785.
- ^ Golovin, A; Henrick (2008). "MSDmotif: zkoumání proteinových stránek a motivů". BMC bioinformatika. 9 (1): 312. doi:10.1186/1471-2105-9-312. PMC 2491636. PMID 18637174.