Skupina ekvivalence - Equivalence group
An ekvivalenční skupina je sada nespecifikovaných buněk, které mají stejný vývojový potenciál nebo schopnost přijímat různé osudy. Náš[SZO? ] současné chápání naznačuje, že skupiny ekvivalence jsou omezeny na buňky stejného původu, známé také jako sourozenecké buňky.[1] Buňky skupiny ekvivalence často přijímají navzájem různé osudy.[2]
Skupiny ekvivalence předpokládají různé potenciální osudy dvěma obecnými, navzájem se nevylučujícími způsoby. Jeden mechanismus, indukce, nastane, když signál pocházející z vnějšku skupiny ekvivalence specifikuje podmnožinu naivních buněk.[2] Další režim, známý jako boční inhibice, vzniká, když signál v rámci skupiny ekvivalence způsobí, že si jedna buňka osvojí dominantní osud, zatímco ostatním ve skupině je bráněno v tom.[3] V mnoha příkladech skupin ekvivalence se k definování vzorů odlišných typů buněk používá indukce i laterální inhibice.
Buňky skupiny ekvivalence, které neobdrží signál, přijmou výchozí osud. Alternativně mohou buňky, které přijímají signál, mít různé osudy.[2][4] V určitém okamžiku se osudy buněk ve skupině ekvivalence nevratně určí, a tak ztratí své multipotentní potenciál. Následující text poskytuje příklady skupin ekvivalence studovaných v hlístice a ascidians.
Skupina ekvivalence prekurzorových buněk vulvy
Úvod
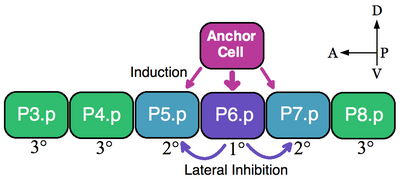
Klasickým příkladem skupiny ekvivalence jsou vulvické prekurzorové buňky (VPC) nematodů. v Caenorhabditis elegans samooplodněná vajíčka opouštějí tělo skrz vulva. Tento orgán se vyvíjí z podskupiny buněk skupiny ekvivalence skládající se ze šesti VPC, P3.p-P8.p, které leží ventrálně podél přední-zadní osy.[5] V tomto příkladu jediná překrývající somatické buňky, kotevní buňka, indukuje blízké VPC, aby získaly vulvální osudy 1 ° (P6.p) a 2 ° (P5.p a P7.p). VPC, které nejsou indukovány, tvoří linii 3 ° (P3.p, P4.p a P8.p), které vytvářejí epidermální buňky, které fúzují na velkou syncyciální epidermis (viz obrázek).[6]
Šest VPC tvoří skupinu ekvivalence, protože všech šest buněk je schopných přijmout jakýkoli z dostupných osudů (1 °, 2 ° a 3 °) v závislosti na jejich blízkosti k kotevní buňce. Ablační experimenty naznačují, že všechny VPC jsou schopny přijmout osud vulvy. Například pokud je buňka P6.p, která se normálně stane 1 °, odstraněna, pak VPC nejblíže kotevní buňce, buď P5.p nebo P7.p, předpokládá osud 1 °. Kromě toho, pokud jsou zničeny všechny VPC kromě nejpřednější buňky P3.p, pak kotevní buňka označuje tuto buňku osud 1 °. Pokud je však kotevní buňka zabita, při absenci indukčního signálu, pak všechny VPC předpokládají výchozí 3 ° linii.[7]
Molekulární mechanismus
Buňka kotvy přímo indukuje osud vulvy vylučováním epidermální růstový faktor (EGF) podobný ligand LIN-3. Buňka P6.p přijímá signál LIN-3 přes receptor tyrosinkinázy LET-23 (P5.p a P7.p také dostávají LIN-3, ale v menší míře). Aktivace LET-23 v P6.p vede k aktivaci LIN-12 (Zářez ) v P5.p a P7.p. Experimentální důkazy ukazují, že LIN-12 je nezbytný a dostatečný pro vytvoření osudu 2 °. Prostřednictvím laterální inhibice LIN-12 brání buňkám P5.p a P7.p přijmout linii 1 °.[7] V tomto příkladu tedy jak skupina induktivní signalizace EGF, tak laterální vzory aktivace Notch skupinu ekvivalence VPC.
Ascidian Pigment Precursor Equivalence Group

Úvod
Larvy ascidianů (mořské stříkačky) obsahují pár senzorických pigmentové buňky známý jako otolit a ocellus. Otolit se používá k snímání gravitace, zatímco ocellus reaguje na světlo. Během embryogeneze se otolit a ocellus vyvíjejí ze dvou bilaterálních ekvivalentních prekurzorů. Buňka prekurzoru levého nebo pravého pigmentu má stejnou pravděpodobnost vývoje v otolit nebo ocellus. Rozhodnutí přijmout jakýkoli osud je určeno po uzavření neurální trubice během rané fáze koncovky (viz obrázek), pomocí špatně definovaného mechanismu indukce.[1]
Během normálního vývoje, po neurální trubice uzávěr, prekurzory pigmentu se dorzálně vyrovnají podél přední-zadní osy neurální trubice. Kterákoli buňka se zarovná vpředu, stane se otolitem, zatímco zadní buňka vytvoří ocellus. Při absenci buněčných interakcí se z obou buněk vyvine ocelli, což je výchozí osud.[3][8]
Experimentální metody studia ekvivalence v systému Windows Halocynthia roretzi
K objasnění, zda jsou osudy otolithu a ocellusu určovány v časném embryu nebo po vyrovnání prekurzorů během uzavření neurální trubice, byly u druhů ascidian použity techniky ablace a léčby drogami Halocythia roretzi.
Buňky, které jsou značeny fluorescein-isothiokyanát-dextranem (FDX), mohou být selektivně fotoabulovány fluorescenční excitací.[9] Když je jedna pigmentová prekurzorová buňka značená FDX fotoabulovaná během stadia střední neurule (15 hodin), z druhé se téměř vždy vyvine ocellus. Pokud jsou však ablace prováděny během pozdní fáze koncového pouzdra (22,5 hodiny), pak zbývající buňka má stejnou pravděpodobnost, že se stane otolitem nebo ocellem.[1]
Inhibice buněčného dělení a morfogeneze s cytochalasin B. je další metoda používaná k určení, kdy je specifikována skupina ekvivalence pigmentového prekurzoru. Léčba cytochalasinem u embryí v časném stadiu (17 hodin), zatímco dvě bilaterální buňky jsou stále odděleny, vede k tomu, že se z obou buněk stane ocelli. Když byl lék použit poté, co se dvě buňky vyrovnaly na hřbetní středové linii, přední buňka se vyvinula do otolitu a zadní buňka se bez výjimky stala ocellus.[1] Oba experimenty naznačují, že osudy prekurzorových buněk pigmentu jsou nevratně určeny přibližně fází středního ocasu (21 hodin).
Další skupiny rovnocennosti
Skupiny ekvivalence byly rovněž popsány v dokumentu ganglion mateřské buňky kobylky a O / P teloblasty v pijavice.[10][11] Stejně jako ostatní případy skupin ekvivalence se buňky potomstva rodí rovnocenné a jsou specifikovány prostřednictvím buněčných interakcí. Skupiny ekvivalence jsou společným tématem ve vývoji mnoha různých organismů phyla.
Reference
- ^ A b C d Nishida a Satoh; Satoh, N (1989). "Stanovení a regulace v linii pigmentových buněk ascidianského embrya". Dev Biol. 132 (2): 355–67. doi:10.1016/0012-1606(89)90232-7. PMID 2494088.
- ^ A b C Greenwald a Rubin; Rubin, GM (1992). „Rozdíl: role interakcí mezi buňkami při vytváření samostatných identit pro ekvivalentní buňky“. Buňka. 68 (2): 271–81. doi:10.1016 / 0092-8674 (92) 90470-W. PMID 1365402.
- ^ A b Nishida; Saitoh, Takashi; Matsumoto, Midori; Makabe, Kazuhiro W .; Nishida, H. (1997). „Notch homolog z Halocynthia roretzi je přednostně exprimován v centrálním nervovém systému během ascidianské embryogeneze.“ Dev Genes Evol. 207 (6): 371–380. doi:10,1007 / s004270050126. PMID 27747436.
- ^ Huang a Weisblat; Weisblat, DA (1996). "Určení osudu buňky ve skupině ekvivalence annelidů". Rozvoj. 122 (6): 1839–47. PMID 8674423.
- ^ Kornfeld (1997). "Vývoj vulvalů u Caenorhabditis elegans". Trendy Genet. 13 (2): 55–61. doi:10.1016 / S0168-9525 (97) 01005-6. PMID 9055606.
- ^ Sternberg a Horvitz; Horvitz, HR (1986). "Tvorba vzoru během vulválního vývoje u C. elegans". Buňka. 44 (5): 761–72. doi:10.1016/0092-8674(86)90842-1. PMID 3753901.
- ^ A b Sternberg, Paul W. (2005). „Vulval development“. WormBook: 1–28. doi:10.1895 / wormbook.1.6.1. PMC 4781130. PMID 18050418.
- ^ Akanuma; et al. (2002). „Signalizace zářezu se podílí na tvorbě nervového systému u embryí ascidian“. Vývojové geny a evoluce. 212 (10): 459–72. doi:10.1007 / s00427-002-0264-x. PMID 12424517.
- ^ Shankland a Weisblat; Weisblat, DA (1984). „Postupné nasazování osudů vysokých buněk během poziční specifikace buněčných linií O a P v embryu pijavice“. Dev Biol. 106 (2): 326–42. doi:10.1016/0012-1606(84)90231-8. PMID 6500176.
- ^ Kuwada a Goodman; Goodman, CS (1985). "Neuronální stanovení během embryonálního vývoje kobylkového nervového systému". Dev Biol. 110 (1): 114–26. doi:10.1016/0012-1606(85)90069-7. PMID 4007260.
- ^ Kuo a Shankland; Shankland, M (2004). „Evoluční diverzifikace specifikačních mechanismů v rámci skupiny O / P ekvivalence pijavice rodu Helobdella“. Rozvoj. 131 (23): 5859–69. doi:10.1242 / dev.01452. PMID 15525668.