Ptačí metaavulavirus 2 - Avian metaavulavirus 2
tento článek potřebuje víc lékařské odkazy pro ověření nebo se příliš spoléhá na primární zdroje. (Červen 2015) |
| Ptačí metaavulavirus 2 | |
|---|---|
| Klasifikace virů | |
| (bez hodnocení): | Virus |
| Oblast: | Riboviria |
| Království: | Orthornavirae |
| Kmen: | Negarnaviricota |
| Třída: | Monjiviricetes |
| Objednat: | Mononegavirales |
| Rodina: | Paramyxoviridae |
| Rod: | Metaavulavirus |
| Druh: | Ptačí metaavulavirus 2 |
| Synonyma | |
Ptačí metaavulavirus 2, dříve Ptačí paramyxovirus 2, je druh viru patřící do čeledi Paramyxoviridae a rod Metaavulavirus. Virus je negativní řetězec RNA virus obsahující a monopartitní genom. Ptačí metaavulavirus 2 je jedním z devíti druhů patřících do rodu Metaavulavirus. Nejčastější sérotyp z Avulavirinae je sérotyp 1, příčina Newcastleská choroba (ND).[Citace je zapotřebí ] Ptačí metaavulavirus 2 je známo, že způsobuje onemocnění, zvláště mírné respirační infekce v domácím drůbež, počítaje v to krůty a kuřata, a má mnoho ekonomických dopadů na produkce vajec a drůbeží průmysl.[5] Virus byl nejprve izolován z kmene v Yucaipa, Kalifornie v roce 1956. Od té doby byly celosvětově izolovány další izoláty viru.

Virová klasifikace
Ptačí metaavulavirus 2 je negativní virus RNA s a monopartitní genom.[5] Patří to Skupina V z Baltimorův klasifikační systém virů.
Struktura
Všechno Paramyxovirus virové částice se skládají z a ribonukleoprotein jádro a lipoproteinová dvojvrstva.[7] Ptačí paramyxoviry obsahují virovou obálku, která zapouzdřuje sférickou virovou kapsidu obsahující genom negativní RNA. Členové Paramyxoviridae rodina, včetně Ptačí metaavulavirus 2postupujte podle „pravidla šesti“, přičemž délka genomu je násobkem šesti. To umožňuje správné zabalení genomu do nukleokapsid.[8] Struktura obalu je charakterizována hrotovitými projekcemi dvou virových proteinů, fúzního proteinu (F) a proteinu hemaglutinin-neuraminidázy (Hn). Tyto proteinové projekce se však mohou u devíti sérotypů ptačích paramyxovirů lišit.[8] Předpokládá se, že protein matrice (M) tvoří vnitřní vrstvu obalu a interaguje s proteiny F a Hn na vnější straně virového obalu během vstupu viru.[8]
Genom
Ptačí metaavulavirus 2 má RNA-gen negativního smyslu, který se po vstupu do buňky musí přepsat do pozitivního smyslu mRNA za účelem produkce virových proteinů a replikace genomu. Bylo předpokládáno elektronová mikroskopie, že genom předpokládá levostrannou spirálovou strukturu.[7] Velikost genomu izolovaného kmene Yucaipa, izolovaného od kuřat v Yucaipě v Kalifornii v roce 1956,[9] je 14 904 nukleotidů dlouhý a obsahuje vedoucí sekvenci 55 nukleotidů na 3 'konci a přívěsnou sekvenci 154 nt na 5' konci.[10] Jiné izolované kmeny Ptačí metaavulavirus 2 byly izolovány, které obsahují genomy podobné, ale odlišné délky. Ptačí metaavulavirus 2 je jedním z nejmenších v Paramyxoviridae rodina.[10] Genom kóduje šest virových proteinů, které jsou od sebe odděleny prostřednictvím intergenové oblasti které jsou mezi 3-23 nukleotidy dlouhé.[10] Šest virových genů je obklopeno oblastmi transkripce zastavte a spusťte signály, které pomáhají při transkripci genomu a produkci subgenomových RNA. Virové genomy většiny sérotypů paramyxovirů kódují geny pro nukleokapsidový protein (N), fosfoprotein (P), matrixový protein (M), fúzní protein (F), hemaglutinin-neuraminidázový (Hn) protein a velký polymerázový protein (L).[8]
Replikační cyklus
Příloha a vstup
Společným znakem všech paramyxovirů je přítomnost dvou membránových kotev glykoproteiny, jeden pro připojení virionu a jeden pro fúzi.[11] Předpokládá se, že proteiny F a Hn přítomné na vnějším obalu viru pomáhají při vstupu viru do hostitelské buňky. Fúzní protein (F) je integrální membránový protein, který sdílí mnoho funkcí podobných jiným virovým fúzním proteinům[11] a je přímo zapojen do fúze virů. Předpokládá se, že dvě oblasti opakování heptadu (HR1) a (HR2) tvoří strukturu svazku šesti šroubovice, která pomáhá indukovat fúzi virové membrány s membránou hostitelské buňky.[12] Protein Hn je tvořen stopkou a kulovitou hlavou, která obsahuje schopnost vázat receptory, které se vážou na receptory kyseliny sialové umístěné na buňkách v horních dýchacích cestách hostitele.[11] Předpokládá se, že interakce mezi Hn a F proteiny způsobují konformační změny v příslušných proteinech, které řídí připojení a fúzi virové kapsidy s membránou hostitelské buňky.[8]
Virová replikace a produkce virového proteinu

Paramyxoviry kódují pouze jednu RNA polymerázu a předpokládá se, že replikace probíhá v cytoplazmě buňky bez jakéhokoli zapojení nebo vstupu do jádra hostitelské buňky.[7] Málo se rozumí tomu, jak polymeráza pomáhá jak při replikaci viru, tak při transkripci a jak lze tyto procesy regulovat.[13] Kódovaná polymeráza se skládá ze dvou proteinů, velké podjednotky odpovědné za syntézu a zakončování RNA a kódovaného proteinu fosfoproteinu (P), který působí jako kofaktor.[13] Jak je popsáno, virový genom zahrnuje intergenní oblasti mezi virovými geny se specifickými start a stop signály. Polymeráza je rozpoznává cis působící prvky ve virovém genomu a iniciuje transkripci.[13] Transkripce nejprve začíná na 3 'konci genomu v délce 40-55 nukleotidů promotor region. Produkuje polymeráza subgenomové RNA transkripcí každého genu, uvolněním produkované RNA a následným vyhledáním dalšího genu pro transkripci cis působícími prvky přítomnými v genomu.[13] Předpokládá se, že signál, při kterém polymeráza uvolňuje transkribovanou RNA jediného genu, obsahuje a Poly U trakt, což vede k omezení a poly (A) ocas k přepsané RNA.[13] Na rozdíl od produkce virové mRNA pro syntézu virových proteinů polymeráza při replikaci virového genomu nezačíná a nezastavuje se na uvedených cis působících prvcích mezi každým genem. Místo toho polymeráza prodlužuje RNA po celé délce genomu. Výše je popsán jeden model pro předpokládanou virovou replikaci a produkci proteinu paramyxovirů, existují však i jiné modely, které naznačují, že polymeráza iniciuje replikaci a transkripci v různých oblastech genomu.[13]
Montáž a uvolnění
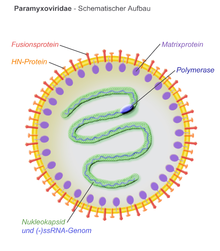
Předpokládá se, že matricový protein (M) na vnitřní straně virového obalu hraje hlavní roli v pučení virových částic z infikované hostitelské buňky.[14] M protein interaguje s Hn a F proteiny, což pomáhá začlenit tyto proteiny do virových částic pro uvolnění.[8] Předpokládala se hypotéza, že matrixové proteiny paramyxovirů interagují se systémem třídění vakuolárního proteinu hostitele, aby zahájily pučení membrány hostitelské buňky. Interakce proteinů Hn, F a M nejsou dobře známy, předpokládá se však, že sestava a uvolňování paramyxoviru závisí na interakcích protein-protein a specifickém cílení proteinů na oblasti hostitelské buňky a membrány[14]
Interakce s hostitelem
V buněčné kultuře mnoho paramyxovirů exprimuje a cytopatický účinek podle syncytium formace, fúze infikovaných buněk se sousedními buňkami produkující velkou vícejadernou buňku.[8] Určité sérotypy ptačího paramyxoviru však nevykazují tvorbu syncytia, pouze zaokrouhlování buněk. Studie prokázaly, že po infekci se mikroskopické léze tvoří v průdušnici, plicích a vzácněji ve střevech zvířecích modelů infikovaných Ptačí metaavulavirus 2.[5] Rovněž bylo prokázáno, že modelové ptáky infikované izoláty z Ptačí metaavulavirus 2 mít průjem a ukázat zvětšený pankreas.[5]
Tropismus
Ptačí metaavulavirus 2 Předpokládá se, že infikuje tkáně dýchacích cest a v některých případech tkáně plic. Virový obalový protein, Hn, se váže na zbytky kyseliny sialové přítomné v buňkách v horních dýchacích cestách. Bylo zjištěno, že u kuřat mohou někteří vykazovat mírné respirační příznaky nebo mohou být asymptomatičtí.[9] Předpokládá se, že u krůt se vyskytují závažnější příznaky včetně zánětu vedlejších nosních dutin a snížení produkce vajec.[9] Byly hlášeny další příznaky, včetně snížené líhnivosti a neplodnosti vajec.[6]
Distribuce virů
Ptačí metaavulavirus 2 má nádrže u divokých druhů ptáků a je známo, že způsobuje onemocnění dýchacích cest a má ekonomické dopady na produkci vajec a drůbežářský průmysl[15] Přirození hostitelé Ptačí metaavulavirus 2 zahrnují krůty a pěvci. Mezi další hostitele však patří kuřata, kolejnice a psittaciny. Ptačí metaavulavirus 2 byl hlášen u ptáků v Evropě, Asii a Severní a Jižní Americe. V USA bylo prokázáno, že virus infikuje krůty s větší pravděpodobností než kuřata.[9] Izoláty z Ptačí metaavulavirus 2 byly získány z oblastí v USA, Bulharsku, Kostarice, Africe a na mnoha dalších místech, což znamená Ptačí metaavulavirus 2 se vyskytuje po celém světě.[5]
Přenos a řízení
K přenosu dochází mezi domestikovanými hejny krůt ve Spojených státech pomalu a způsob přenosu není dobře definován.[6] Předpokládá se, že infekce Ptačí metaavulavirus 2 u domestikované drůbeže pochází z interakcí s infikovanými divokými ptáky.[6] Je možné přijmout preventivní opatření, aby se zabránilo šíření Ptačí metaavulavirus 2 v domestikovaných hejnech. Mezi tato preventivní opatření patří minimalizace interakcí mezi hejny a volně žijícími ptáky v oblastech odolných proti ptákům, kde je chována a chována drůbež, a správné hygienické postupy.[6] Nebyly hlášeny žádné lidské infekce Ptačí metaavulavirus 2[6]
Reference
- ^ Balkema-Buschmann, Anne; et al. (6. června 2018). „Reorganizace rodiny Paramyxoviridae". Mezinárodní výbor pro taxonomii virů (ICTV). Citováno 3. května 2019.
- ^ Rima, Bertus K .; et al. (Červen 2016). "Přidání jednoho (1) nového druhu a přejmenování dvanácti (12) druhů rodu Avulavirus" (PDF). Mezinárodní výbor pro taxonomii virů (ICTV). Citováno 3. května 2019.
Chcete-li přejmenovat následující taxon (nebo taxony): Názvy druhů v rodu Avulavirus. Aktuální název Navrhovaný název Ptačí paramyxovirus 2 Ptačí avulavirus 2
- ^ 3. zpráva ICTV Matthews, R. E. F. (1979). Klasifikace a nomenklatura virů. Třetí zpráva Mezinárodního výboru pro taxonomii virů. Intervirology 12: 129-296. p217 https://talk.ictvonline.org/ictv/proposals/ICTV%203rd%20Report.pdf
- ^ 1 ICTV 5th Report Francki, R. I. B., Fauquet, C. M., Knudson, D. L. & Brown, F. (eds) (1991). Klasifikace a nomenklatura virů. Pátá zpráva Mezinárodního výboru pro taxonomii virů. Archives of Virology Supplementum 2, p244 https://talk.ictvonline.org/ictv/proposals/ICTV%205th%20Report.pdf
- ^ A b C d E Warke, Ashwini; Stallknecht, David; Williams, Susan M .; Pritchard, Nikki; Mundt, Egbert (2008). „Srovnávací studie o patogenitě a imunogenicitě izolátů volně žijících ptáků ptačího paramyxoviru 2, 4 a 6 u kuřat“. Ptačí patologie. 37 (4): 429–434. doi:10.1080/03079450802216645. PMID 18622861. S2CID 3995370.
- ^ A b C d E F „Jiné ptačí infekce paramyxovirem - drůbež - Veterinární příručka Merck“. Veterinární příručka Merck. Citováno 2017-11-02.
- ^ A b C Anderson, Danielle (2011). Biologie paramyxovirů. ISBN 9781904455851.
- ^ A b C d E F G Kim, Shin-Hee (2012). „Replikace, neurotropismus a patogenita ptačích paramyxovirových sérotypů 1-9 u kuřat a kachen“. PLOS ONE. 7 (4): e34927. Bibcode:2012PLoSO ... 734927K. doi:10.1371 / journal.pone.0034927. PMC 3340391. PMID 22558104.
- ^ A b C d Zarkov, Iv (2015). "Přehled infekce ptačími paramyxovirovými sérotypy 2 (APMV-2) a první výsledky Bulharska" (PDF).
- ^ A b C Subbiah, Madhuri; Xiao, Sa; Collins, Peter L .; Samal, Siba K. (01.10.2008). "Kompletní sekvence genomu ptačího paramyxoviru typu 2 (kmen Yucaipa) a srovnání s jinými paramyxoviry". Virový výzkum. 137 (1): 40–48. doi:10.1016 / j.virusres.2008.05.012. PMC 2700353. PMID 18603323.
- ^ A b C „Položka Paramyxovirus (stažení PDF k dispozici)“. ResearchGate. Citováno 2017-11-02.
- ^ Wang, Xiao-Jia; Bai, Ya-Duo; Zhang, Guo-Zhong; Zhao, Ji-Xun; Wang, Ming; Gao, George F. (duben 2005). "Struktura a funkce studie paramyxovirového fúzního proteinu heptad opakujících se peptidů". Archivy biochemie a biofyziky. 436 (2): 316–322. doi:10.1016 / j.abb.2005.02.004. PMC 7094221. PMID 15797244.
- ^ A b C d E F Noton, Sarah L .; Fearns, Rachel (01.05.2015). "Zahájení a regulace transkripce a replikace paramyxoviru". Virologie. Vydání k 60. výročí. 479-480 (dodatek C): 545–554. doi:10.1016 / j.virol.2015.01.014. PMC 4424093. PMID 25683441.
- ^ A b Pantua, Homer D .; McGinnes, Lori W .; Peeples, Mark E .; Morrison, Trudy G. (01.11.2006). „Požadavky na shromáždění a uvolnění částic podobných viru newcastleské choroby“. Journal of Virology. 80 (22): 11062–11073. doi:10.1128 / JVI.00726-06. ISSN 0022-538X. PMC 1642154. PMID 16971425.
- ^ Alexander, D.J (2000). „Newcastleská choroba a jiné ptačí paramyxoviry“ (PDF). Revue Scientifique et Technique (International Office of Epizootics). 19 (2): 443–62. PMID 10935273.