Tatuidris - Tatuidris
| Tatuidris | |
|---|---|
 | |
| T. tatusia pracovník z El Campano, Magdaléna, Kolumbie[1] | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Arthropoda |
| Třída: | Insecta |
| Objednat: | Blanokřídlí |
| Rodina: | Formicidae |
| Podčeleď: | Agroecomyrmecinae |
| Rod: | Tatuidris Hnědá & Kempf, 1968 |
| Druh: | T. tatusia |
| Binomické jméno | |
| Tatuidris tatusia Brown & Kempf, 1968 | |
| Synonyma | |
T. kapasi Lacau & Groc, 2012 | |
Tatuidrisnebo pásovec ant, je vzácný rod z mravenci skládající se z jediného druh, Tatuidris tatusia. Mravenci jsou malé velikosti a obývají podestýlka z Neotropické lesy v Centrální a Jižní Amerika, z Mexiko na Brazílie. Pracovníci mají železnou barvu až tmavě červenou barvu a jsou charakteristické morfologie, skládající se z hlavy podobné štítu se širokým vrchol, ventrálně těžký čelisti které se při úplném uzavření nepřekrývají a mezi mravenci jedinečné - an anténa zásuvkový aparát sedí vzhůru nohama. O biologii mravenců je známo jen málo, ale je pravděpodobné noční a specialista predátoři.
Tatuidris byl poprvé popsán v roce 1968 a původně umístěn do myrmicin kmen Agroecomyrmecini spolu se dvěma fosilními rody. Od původního popisu byla debata zaměřena na systematický stav kmene.
Taxonomie
Tatuidris tatusia je jediným druhem v Tatuidris, a monotypický rod a jeden z pouhých dvou existujících rodů v podčeledi Agroecomyrmecinae.[2][3] Nový druh, T. kapasi, popsali Lacau & Groc v roce 2012,[4] ale nyní byl zařazen do a junior synonymum pod T. tatusia na základě rozsahu morfologické variability vyskytující se v celém tomto širokém geografickém rozsahu. Analýza Čárové kódy DNA označil vzor genetická izolace podle vzdálenosti, což naznačuje přítomnost jediného podstupovaného druhu alopatrická diferenciace.[5]
Poprvé to popsali Brown & Kempf v roce 1968 na základě dvou pracovníků shromážděných v a Berlese vzorek z humus v El Salvador.[6][7] Kvůli morfologickým podobnostem to považovali za velmi primitivní mravenec a umístil jej do tehdejšího a myrmicin kmen, Agroecomyrmecini, spolu s mravenci známými z Raný eocén Baltský jantar (Agroecomyrmex ) a pozdní eocén Florissantská břidlice (Eulithomyrmex ).[7]
Má povrchní podobnost s některými existujícími rody (Strumigenys, Ishakidris, Pilotrochus, a Phalacromyrmex ), ale tyto podobnosti jsou považovány za způsobené konvergentní evoluce. Kvůli podobnostem v habitu, Brown & Kempf (1968) spojili Tatuidris do Dacetini rod Glamyromyrmex (v současné době juniorské synonymum pro Strumigenys) a Phalacromyrmex. Došli však k závěru: „analýza těchto podobností naznačuje [...], že jsou většinou konvergentní a nejsou založeny na blízkém fylogenetickém vztahu“.[8] Další práce zkoumaly podobnosti Tatuidris s Ishakidris (Bolton 1984) a Pilotrochus (Brown 1977). Zatímco tyto taxony sdílejí některé vlastnosti, včetně rozšířeného vrcholu hlavy, hlubokých anténních scrobů a kompaktu mezosoma, podobnosti byly opět považovány za konvergentní.[5][9][10]


Od původního popisu byla debata zaměřena na systematický stav kmene. Bolton (2003) jako první navrhl taxonomickou nestabilitu Tatuidris v Myrmicinae a pozvedl rod na úroveň nové podčeledi, Agroecomyrmecinae, což naznačuje, že Agroecomyrmecinae může být sesterským taxonem Myrmicinae.[12][13] Hodnost podčeledi byla znovu posouzena Baroni Urbani & de Andrade v roce 2007, toto byl první pokus zahrnout Tatuidris jako terminální taxon v morfologické kladistické analýze. Ve své studii Baroni Urbani & de Andrade identifikovali morfologické znaky synapomorfie sdílené mezi Tatuidris a dacetiny, které ospravedlňují zahrnutí rodu do Myrmicinae.[14][15] Navíc dva autapomorphies (řapík různě tvarovaný tergum a hrudní kost a oči na vrcholu anténního scrobu nebo v jeho blízkosti) oddělené Tatuidris ze všech ostatních existujících rodů mravenců zahrnutých do jejich studie.[15][16]
Na rozdíl od fylogenetické studie založené na morfologických vlastnostech, molekulární analýzy vnitřní fylogeneze mravenců poskytly silné důkazy o tom, že mravenci pásovci nejsou úzce spjati s Myrmicinae ani se do nich nevnořují. Brady et al. (2006), Moreau et al. (2006) a Rabeling et al. (2008) rekonstruovali fylogenetické stromy agroecomyrmeciny uvnitř „poneroidní“ skupiny podrodin, v blízkosti Paraponerinae, a podpořilo vyloučení rodu z Myrmicinae, podčeledi umístěné uvnitř „formicoidního“ kladu.[17] Vzhledem k časnému výskytu Agroecomyrmecinae v geologickém záznamu se předpokládalo, že podobnosti mravenců pásovce s Myrmicinae představují konvergenci a / nebo retenci plesiomorfní formuláře.[18][19]
Název Tatuidris znamená „pásovec mravenec ", což je také běžné jméno pro tento druh;[20] tatu pochází z Tupi a portugalské slovo pro „pásovce“; the konkrétní epiteton pro jednotlivé popsané druhy, tatusie, je starý obecný název pro pásovce.[6]
Rozdělení
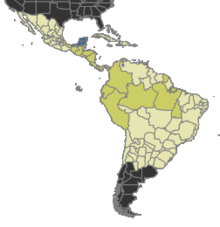
Současnost, dárek | Pravděpodobně přítomný |
Nejistý | Nepřítomen |
Tatuidris je vzácný, ale široce distribuovaný.[19] Mravenci obývají vrh listů Neotropické lesy ve Střední a Jižní Americe, od Mexika po Francouzská Guyana,[21] centrální Brazílie,[22] a Amazonské Peru.[5] Z kolekce nejsou známy žádné sbírky karibský, Galapágy nebo jiné ostrovy. V současnosti je známo, že většina vzorků a sbírek se vyskytuje v lokalitách západně od Andy, přičemž více sbírek se obvykle vyskytuje ve Střední Americe a Mexiku. Většina sbírek pochází z horských (předhorských) oblastí ve středních nadmořských výškách (obvykle 800–1200 metrů nadmořské výšky). Sbírky z nížiny Amazonský deštný prales je málo.[23] Publikovaných záznamů je málo, ale s příchodem prosévání vrhů a Winklerova extrakce jako oblíbená metoda sběru mravenců, Tatuidris nejsou tak vzácné, jak se zdálo. Ačkoli to není příliš hojné, s častým proséváním vrhu je lze spolehlivě najít Kostarické mokré lesy.[7]
Popis
Pracovníci Tatuidris představují výraznou morfologii, skládající se z hlavy podobné štítu se širokým vrcholem (horní povrch hlavy), ventrálně otočených těžkých čelistí, které se při úplném uzavření nepřekrývají, hlubokých anténních scrobů (dojem, který přijímá části antény) s očima na vrcholu nebo blízko jejich vrcholu, kompaktní a fúzovaný mezosom, 7 segmentová anténa, první gastrální segment ventrálně nasměrovaný a mezi mravenci jedinečný - zásuvkový anténní aparát sedící vzhůru nohama na střeše rozšířeného čelního laloku.[5][24]
Tělo pracovníka je krátké a kompaktní, železný -barevné až tmavě červené, silné a tuhé kůže (vnější "kůže"). Tělo je pokryto chloupky, které se liší délkou a sklonem. Hlava je pyriformní (hruškovitého tvaru), nejširší zezadu, s malými očima. Královny jsou podobné pracovníkům, ale žlutě zbarvené a bledší. Jejich křídla jsou asi o 60% delší než celková délka těla. Muži, rovněž podobní pracovníkům (kromě hlavy), jsou tmavší a mají křídla asi o 50% delší než tělo. Oči královen a mužů jsou větší než u dělníků.[25]
Velikost
Exempláře Tatuidris jsou malé, asi 3,5 mm (0,14 palce)[A] v celkové délce, ale vzorky se mohou značně lišit ve velikosti, přičemž větší vzorky jsou dvakrát větší než ty menší.[b] Variabilita velikosti úlovků pastí (možná stejných kolonií) může být značná. Například se chytí pracovníci z jedné sbírky Nikaragua měnil 30% velikosti. Stále není jasné, zda je kolísání velikosti kolonií způsobeno přítomností morfologických kast pracovníků (např. Malých a velkých kast) nebo kontinuální variabilitou velikosti. Analýza hlavních komponent (PCA) odhalilo, že většina variability mezi vzorky souvisí s velikostí, přičemž tvar vysvětluje jen malou část celkové variace.[27]
Variabilita filozofie

Čtyři pilositické vzory (vzory podobné vlasům štítek ) je známo, že se vyskytují uvnitř Tatuidris sbírky. Filosofický vzor A se skládá ze směsi dlouhých flexuózních i krátkých appressovaných štětin. Toto je nejběžnější vzor pilosity a ten, který se nejvíce podobá typovým vzorkům ze Salvadoru a gyne z Ekvádoru v Otongachi. Filosofický vzor B je charakterizován velmi krátkými, plně stlačenými a pravidelně rozmístěnými štětinami uspořádanými homogenně a ve stejné vzdálenosti na hlavě, mesosoma, řapík, postpetiole a gaster. Filosofický vzor C je charakterizován hustými štětinami vypadajícími jako lanosa. Filosofický vzor D se skládá z krátkých a stejnoměrných polohlavých (silně nakloněných, ale ne zcela potlačených) štětin rozptýlených po celém těle.[28]
Oči
Relativní poloha očí je v rámci druhu velmi variabilní. Například umístění očí se pohybuje od toho, že je úplně v anténních scrobech, až po úplně mimo scroby. V některých případech je samotné oko umístěno mimo anténní scrob, ale fossa oka je dobře označena a splývá s anténním scrobem. Ve většině vzorků se anténní carina (hřeben táhnoucí se podél hřbetní anténní oblasti) rozdvojuje z anténních šupin a leží přímo nad očima. Ve vzorcích z Nikaraguy je však přítomna silně ohromená anténní carina. U těchto vzorků leží asi 40% oblasti oka v anténních šupinách. V gynekologii leží pouze ~ 1/6 oka v anténních šupinách. Deprese se někdy tvoří v kůži po stranách propodeum, pod propodeal průduch a nad metapleurální žlázou. Hloubka této deprese se u jednotlivých vzorků liší a u větších vzorků má tendenci být nejhlubší.[29]
Biologie
Biologie je málo známá; mužské a ženské reprodukční kasty byly popsány poprvé v roce 2012.[19] Rod je znám hlavně z izolovaných pracovníků nalezených ve Winkleru nebo Berlese Vzorky.[7] Tatuidris pracovníci mají zvláštní mandibulární štětce a silné bodnutí, což vedlo společnost Brown & Kempf ke spekulaci Tatuidris mohou být speciálními predátory aktivních nebo kluzkých členovec kořist.[6] Do roku 2011 nebyla registrována žádná pozorování živých vzorků.[30] Podrobnosti o první kolekci malého živého přenosu kolonie (3 pracovníci a 4 gyny) od Thibauta Delsinneho v lesích střední výšky na jihovýchodě Ekvádor navrhuji, aby Tatuidris může být vysoce specializovaným predátorem, protože kolonie chované v zajetí nepřijímaly žádnou potravinu, která jim byla nabídnuta. Potraviny odmítané mravenci zahrnovaly živé i mrtvé termiti, mnohonožky, roztoči, různé části hmyzu, cukrová voda, tuňák, sušenky, živé a mrtvé ovocné mušky (Drosophila ), žít jarní chvilky, žít myriapods (Chilopoda a Diplopoda ), živí a mrtví Diplura, malé živé pavouky, malé živé pseudoscorpions, jeden malý hlemýžď, nevařené slepičí vejce (tj. kousek vaty namočený do čerstvého rozšlehaného slepičího vejce), larvy mravenců (Gnamptogenys sp.) a živých mravenců (Cyphomyrmex sp., Brachymyrmex sp.). Potenciální potravinové položky (členovci) pro Tatuidris byly odebrány ze vzorků půdy a Winklerových vzorků shromážděných na místě, kde Tatuidris byl a priori rozhodnut být přítomen.[23]
Další pozorování tomu nasvědčují Tatuidris může být dravec sedět a čekat.[31][32] Delsinne poznamenal, že „jak pracovníci, tak gynes pohyboval se velmi pomalu a byl velmi nemotorný. Často zůstali nehybní během několika desítek sekund nebo dokonce několika minut, když byli vyrušeni (ať už mým zacházením nebo kontaktem s jiným členovcem). “[33] Tato pozorování byla prováděna hlavně v noci, což tomu nasvědčuje Tatuidris může být noční, hypotéza také podporována vzory sběru. Například v ekvádorském lese Río Toachi Tatuidris vzorky mají tendenci spadat nástrahy, místo Winklerových vaků.[34] Vzhledem k tomu, že nástrahové pasti obvykle fungují 24 hodin, ale Winklerovy vaky obecně používají stelivo prosívané během dne, pak mohou být ve vzorcích Winklera mravenci s nočními návyky nedostatečně zastoupeni. Malé oči Tatudris druhy poskytují další podporu pro tuto hypotézu.[33]
Poznámky
- ^ Formálně bylo popsáno několik vzorků. Celková délka (včetně čelistí) T. tatusia pracovník holotypu je 3,5 mm a vzorek, který byl použit k popisu T. kapasi je 3,42 mm.[21][26]
- ^ Na základě vzorků naměřených Donosem (2012), průměr Weberova délka (v bočním pohledu, od základny předního sklonu pronotum po spodní posteroventrální úhel propodea) je 0,62 mm, s minimem 0,45 mm a maximem 0,90 mm.[27]
Reference
- ^ „Vzor: PSW7891-9 Tatuidris tatusia". antweb.org. AntWeb. Citováno 3. září 2013.
- ^ Bolton, B. (2015). "Tatuidris". AntCat. Citováno 13. listopadu 2015.
- ^ Bolton, B. (2015). "Agroecomyrmecinae". AntCat. Citováno 13. listopadu 2015.
- ^ Lacau a kol. 2012, s. 1–6
- ^ A b C d Donoso 2012, str. 61
- ^ A b C d Brown & Kempf 1968, str. 189
- ^ A b C d "Rod: Tatuidris". antweb.org. AntWeb. Citováno 29. srpna 2013.
- ^ Brown & Kempf 1968, str. 183
- ^ Bolton 1984, str. 380
- ^ Brown 1977, str. 222
- ^ "Vzor: CASENT0178755 Tatuidris tatusia". antweb.org. AntWeb. Citováno 2. září 2013.
- ^ Bolton 2003, str. 51
- ^ Donoso 2012, s. 61–62
- ^ Baroni Urbani & de Andrade 2007, str. 78
- ^ A b Donoso 2012, str. 62
- ^ Baroni Urbani & de Andrade 2007, str. 80–81
- ^ Oddělení 2007, str. 555–557
- ^ Oddělení 2011, str. 23
- ^ A b C Donoso 2012, str. 63
- ^ Lacau a kol. 2012, str. 1
- ^ A b Lacau a kol. 2012, str. 4
- ^ Vasconcelos & Vilhena 2002, str. 278
- ^ A b Donoso 2012, str. 71
- ^ Keller 2011, str. 27
- ^ Donoso 2012, s. 67–68
- ^ Brown & Kempf 1968, str. 187
- ^ A b Donoso 2012, str. 70
- ^ Donoso 2012, str. 69
- ^ Donoso 2012, s. 72–73
- ^ Jacquemin a kol. 2014, str. 4
- ^ Jacquemin a kol. 2014, str. 5
- ^ Billen & Delsinne 2013, str. 63
- ^ A b Donoso 2012, str. 72
- ^ Donoso & Ramón 2009, str. 491
- Baroni Urbani, C .; de Andrade, M.L. (2007), „Kmen mravenců Dacetini: Meze a základní rody, s popisem nových druhů“, Annali del Museo Civico di Storia Naturale "G. Doria", 99: 1–191
- Billen, Johan; Delsinne, Thibaut (2013), „Nová intramandibulární žláza u mravence Tatuidris tatusia (Hymenoptera: Formicidae) ", Myrmekologické zprávy, 19: 61–64
- Bolton, B. (1984), „Diagnóza a vztahy rodu myrmicinů mravenců Ishakidris gen. N. (Hymenoptera: Formicidae).“, Systematická entomologie, 9 (4): 373–382, doi:10.1111 / j.1365-3113.1984.tb00516.x
- Bolton, B. (2003), Synopse a klasifikace Formicidae, Monografie amerického entomologického institutu, 71, The American Entomological Institute, str. 1–370
- Brady, S.G .; Schultz, T.R .; Fisher, B.L .; Ward, P.S. (2006), „Hodnocení alternativních hypotéz pro časný vývoj a diverzifikaci mravenců“, Sborník Národní akademie věd, 103 (48): 18172–18177, Bibcode:2006PNAS..10318172B, doi:10.1073 / pnas.0605858103, PMC 1838725, PMID 17079492
- Brown, W.L. (1977), „Aberrant new rod of myrmicine ant from Madagascar“, Psychika, 84 (3–4): 218–224, doi:10.1155/1977/41590
- Brown, W. L., Jr.; Kempf, W. W. (1968), "Tatuidris, pozoruhodný nový rod Formicidae (Hymenoptera) ", Psychika (publikováno 1967), 74 (3): 183–190, doi:10.1155/1967/868759
- Donoso, D.A. (2012), „Dodatky k taxonomii mravenců pásovce (Hymenoptera, Formicidae, Tatuidris)" (PDF), Zootaxa, 3503: 61–81, doi:10.11646 / zootaxa.3503.1.5
- Donoso, D.A .; Ramón, G. (2009), „Složení vysoce rozmanité společenstva mravenců (Hymenoptera: Formicidae) z ekvádorského předhorského deštného pralesa“, Annales de la Société Entomologique de France, 45 (4): 487–499, doi:10.1080/00379271.2009.10697631, S2CID 83979652
- Jacquemin, J .; Delsinne, T .; Maraun, M .; Leponce, M. (2014), „Trofická ekologie mravence pásovce, Tatuidris tatusia, hodnoceno stabilními izotopy a pozorováním chování ", Journal of Insect Science, 14 (108): 1–12, doi:10.1673/031.014.108, PMC 4700978, PMID 25199767
- Keller, R.A. (2011), „Fylogenetická analýza morfologie mravenců (Hymenoptera: Formicidae) se zvláštním odkazem na podrodiny poneromorfů“, Bulletin of the American Museum of Natural History, 355: 1–90, doi:10.1206/355.1, hdl:2246/6124, S2CID 84005152
- Lacau, Sébastien; Groc, Sarah; Dejean, Alain; Oliveira, Muriel L. de; Delabie, Jacques H. C. (2012), "Tatuidris kapasi sp. listopad: nový mravenec pásovec z Francouzské Guyany (Formicidae: Agroecomyrmecinae). ", Psychika, 2012: 1–6, doi:10.1155/2012/926089
- Moreau, C.S .; Bell, C.D .; Vila, R .; Archibald, S.B .; Pierce, N.E (2006), „Fylogeneze mravenců: diverzifikace ve věku krytosemenných rostlin“, Věda, 312 (5770): 101–104, Bibcode:2006Sci ... 312..101M, doi:10.1126 / science.1124891, PMID 16601190, S2CID 20729380
- Rabeling, C .; Brown, J.M .; Verhaagh, M. (2008), „Nově objevená sesterská linie vrhá světlo na časnou evoluci mravenců“, Sborník Národní akademie věd, 105 (39): 14913–14917, Bibcode:2008PNAS..10514913R, doi:10.1073 / pnas.0806187105, PMC 2567467, PMID 18794530
- Vasconcelos, Heraldo L .; Vilhena, José M.S. (2002), „První záznam o rodu mravenců Tatuidris (Hymenoptera: Formicidae) v Brazílii " (PDF), Revista de Biología Tropical, 51 (1): 278
- Ward, P.S (2007), „Fylogeneze, klasifikace a druhová taxonomie mravenců (Hymenoptera: Formicidae)“, Zootaxa, 1668: 549–563, doi:10.11646 / zootaxa.1668.1.26
- Ward, P.S (2011), „Integrace molekulárních fylogenetických výsledků do taxonomie mravenců (Hymenoptera: Formicidae)“, Myrmekologické zprávy, 15: 21–29
 Tento článek včlení text publikovaný pod Creative Commons Attribution 3.0 Unported License: Donoso, D.A. (2012). „Dodatky k taxonomii mravenců pásovce (Hymenoptera, Formicidae, Tatuidris)" (PDF). Zootaxa. 3503: 61–81. doi:10.11646 / zootaxa.3503.1.5..
Tento článek včlení text publikovaný pod Creative Commons Attribution 3.0 Unported License: Donoso, D.A. (2012). „Dodatky k taxonomii mravenců pásovce (Hymenoptera, Formicidae, Tatuidris)" (PDF). Zootaxa. 3503: 61–81. doi:10.11646 / zootaxa.3503.1.5..- Tento článek včlení text publikovaný pod Creative Commons Attribution 3.0 Unported License: "Rod: Tatuidris". antweb.org. AntWeb. Citováno 29. srpna 2013..
externí odkazy
 Média související s Tatuidris tatusia na Wikimedia Commons
Média související s Tatuidris tatusia na Wikimedia Commons