Celulární dewetting - Cellular dewetting
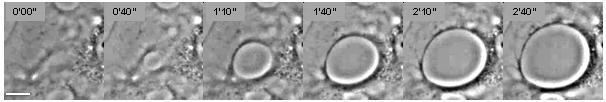
Buněčný dewetting odkazuje na proces nukleace a rozšíření tunelů makroapertury transendoteliálních buněk (TEM) v Liberci endoteliální buňky (Obrázek 1).[1] Tento jev je analogický s nukleací a růstem suchých skvrn ve viskózních kapalinách šířících se na a nezmáčitelné substrát (obrázek 2).[2] Odvlhčování buněk spouští několik proteinových toxinů z patogenní bakterie, zejména faktory podobné EDIN z Zlatý stafylokok a od Clostridium botulinum, stejně jako edémový toxin z Bacillus anthracis.[3][4] TEM se tvoří v reakci na prasknutí cytoskelet fyzické spojení prostřednictvím cytoplazma v důsledku inhibice Dráha RhoA / ROCK nebo k indukci toku cyklického AMP (tábor ) široký signální molekula.[4][5]
Fyzika za buněčným odvlhčováním

Fenomén buněčného odvlhčování lze interpretovat fyzikálním modelováním (obrázek 2).[6] The hnací síla zodpovědný za spontánní tvorbu TEM tunelů a jejich otevření je membrána napětí který je výsledkem šíření buněk v důsledku relaxace aktomyosinu. Na rozdíl od kapalného odvlhčování dosahují TEM maximálního průměru, při kterém je hnací síla vyvážena odporovou silou, která se vyvíjí podél okrajů TEM (obrázek 2). Tato odporová síla se označuje jako napětí vedení a je na molekulární úrovni necharakterizovaný.
Fyzikální parametry
Hnací síly vtažením do tunelu o poloměru R, jak je znázorněno na obrázku 2. Zde je tahání způsobeno napínáním buněčná membrána (σ), které je částečně potlačeno napětím vedení kolem tunelu (T). Za těchto podmínek je čistá hnací síla (FD) se skládá ze dvou příspěvků:

Dewetting pokračuje, pokud FD>0.
Napětí membrány (σ) závisí na poloměru tunelu R. Zvětšení tunelu uvolňuje membrána, vyvolávající pokles membránového napětí, jak je popsáno v Helfrichův zákon.
Napětí vedení (T) odpovídá odporové síle podél okraje tunelu, která je v rozporu s napětím membrány a omezuje odvodnění. Toto napětí linie může mít fyzické a molekulární složky.
Reference
- ^ Lemichez, E. (2012). „Dynamika transcelulárního tunelu: Řízení buněčného odvlhčování kontraktilitou aktomyosinu a proteiny I-BAR“. Biologie buňky. 105 (3): 109–117. doi:10.1111 / boc.201200063. PMID 23189935.
- ^ De Gennes, P.-G. (2004). Fenomény kapilárnosti a zvlhčování. New York: Springer. ISBN 978-0387005928.
- ^ Boyer, L. (2006). "Indukce přechodných makroapertur v endoteliálních buňkách prostřednictvím inhibice RhoA faktory Staphylococcus aureus". Journal of Cell Biology. 173 (5): 809–819. doi:10.1083 / jcb.200509009. PMC 2063895. PMID 16754962.
- ^ A b Maddugoda, M. P. (2011). „Signalizace cAMP toxinem edému antraxu indukuje tunely transendoteliálních buněk, které jsou znovu uzavřeny MIM prostřednictvím polymerace aktinů řízené Arp2 / 3“. Mobilní hostitel a mikrob. 10 (5): 464–474. doi:10.1016 / j.chom.2011.09.014. PMID 22100162.
- ^ Cai, Y. (2010). „Cytoskeletální koherence vyžaduje kontraktilitu myosinu-IIA“. Journal of Cell Science. 123 (3): 413–423. doi:10.1242 / jcs.058297. PMC 2816186. PMID 20067993.
- ^ Gonzalez-Rodriguez, D. (2012). "Buněčné odvlhčení: otevření makroapertur v endotelových buňkách". Dopisy o fyzické kontrole. 108 (21): 218105. doi:10.1103 / PhysRevLett.108.218105. PMID 23003307.