Dělení biomasy - Biomass partitioning
Dělení biomasy je proces, kterým rostliny rozdělit jejich energii mezi jejich listy, stonky, kořeny, a reprodukční části. Tyto čtyři hlavní součásti rostliny mají důležité morfologické role: listy berou v CO2 a energii ze slunce k tvorbě sloučeniny uhlíku, stonky rostou nad konkurenty, aby dosáhly slunečního světla, kořeny absorbují vodu a minerální živiny z půdy při ukotvení rostliny a reprodukční části usnadňují pokračování druhů. Rozdělení rostlin biomasa v reakci na limity nebo přebytky zdrojů, jako je sluneční světlo, oxid uhličitý, minerální živiny a voda, je růst regulován konstantní rovnováhou mezi rozdělením biomasy mezi části rostlin. Dochází k rovnováze mezi růstem kořenů a výhonků, protože kořeny potřebují sloučeniny uhlíku z fotosyntézy ve výhoncích a výhonky potřebují dusík absorbovaný z půdy kořeny.[1] Přidělování biomasy je omezeno na růst; limit pod zemí zaostří biomasu na kořeny a limit nad zemí zvýhodní větší růst výhonku.[2]
Rostliny fotosyntetizovat vytvářet sloučeniny uhlíku pro růst a skladování energie. Cukry vytvořené fotosyntézou jsou poté transportovány phloem za použití průtok tlaku systému a jsou používány pro růst nebo uloženy pro pozdější použití. Rozdělení biomasy způsobí, že se tento cukr rozdělí způsobem, který maximalizuje růst, poskytuje maximální kondici a umožňuje úspěšnou reprodukci. Rostlinné hormony hrají velkou roli při dělení biomasy, protože ovlivňují diferenciaci a růst buněk a tkání změnou exprese genů a změnou morfologie.[3] Reagováním na životní prostředí podněty a odpovídajícím způsobem rozdělit biomasu jsou rostliny schopny lépe přijímat zdroje ze svého prostředí a maximalizovat růst.
Abiotické faktory dělení

Je důležité, aby rostliny dokázaly vyvážit absorpci a využití dostupných zdrojů a upravily svůj růst, aby získaly více vzácných zdrojů omezujících růst: sluneční světlo, oxid uhličitý, minerální živiny a vodu.[4] Rovnováhu dělení biomasy lze vysvětlit Liebigův zákon minima a modelovány prostřednictvím metafory Liebigova hlaveň ve kterém omezující zdroj růstu rostlin je jako nejkratší lišta na sudu plném vody.[5] Sud může zadržovat vodu pouze do úrovně nejkratší lišty a podobně mohou rostliny růst pouze takovou rychlostí, jakou umožňuje omezující zdroj. Aby bylo možné pokračovat v růstu, musí být biomasa rozdělena, aby pomohla tyto zdroje izolovat.
Sluneční světlo
Hlavním mechanismem snímání světla pro rostliny je fytochrom systém s pigmenty rozmístěnými po celé rostlině k detekci změn v červeném a daleko červeném světle.[2] Detekce kvality světla pomocí fytochromů pomáhá spouštět změny v dělení biomasy. Bylo prokázáno, že rostliny pěstované za špatných světelných podmínek přidělují více biomasy výhonkům (hlavně listím). Měřením listové plochy rostlin na různých úrovních ozáření nebo slunečního záření bylo zjištěno, že nižší úrovně světla způsobují zvětšení celkového povrchu listů rostlin.[6] Pokud sluneční světlo omezuje, rostlina zvyšuje růst výhonku a snižuje energii rozdělenou do kořenů, protože nižší rychlost fotosyntézy snižuje potřebu vody a minerálních živin.[2] Protože slabé světlo způsobuje větší alokaci výhonku, je u rostlin ve vysokém světle provedena stejná korelace; vyšší ozáření způsobuje menší celkovou nižší povrchovou plochu, protože k absorpci slunečního záření je potřeba menší povrchová plocha. Když má rostlina více než dostatečnou fotosyntetickou kapacitu, bude místo toho upřednostňovat růst v kořenech, aby absorbovala vodu a živiny.
Oxid uhličitý
V situaci, kdy uhlík je omezujícím zdrojem, zvyšuje hladinu CO2 zvyšuje rychlost fotosyntézy.[7] To také způsobí zvýšení příjmu živin a spotřeby vody se zaměřením většího růstu směrem ke kořenům. Při nízkém CO2 koncentrace, rostliny vytvářejí větší a početnější listy, aby přinesly více CO2.[7] Dopady koncentrace oxidu uhličitého v atmosféře na dělení biomasy je důležité pochopit při dopadech na rostliny tváří v tvář změně klimatu. Rostlinné buňky zvýšily poměr uhlíku k dusíku, když CO2 koncentrace jsou vyšší, což snižuje hladinu rozkladu. Snížení rozkladu způsobené vysokými koncentracemi oxidu uhličitého má za následek snížení dostupnosti dusíku pro rostliny.[8]

Minerální živiny
Celkově má dostupnost živin silný účinek na dělení, přičemž rostliny rostoucí v oblastech se špatnými živinami dělí většinu své biomasy na podzemní struktury.[9] Například dostupnost půdního dusíku je silným determinantem přidělování biomasy. Například v systémech s nízkou produktivitou (nízké hladiny dusíku v půdě), jako jsou boreální lesy, stromy věnují velkou část své biomasy kořenům. Ale jak se zvyšuje produktivita půdy, biomasa se primárně přiděluje nadzemním strukturám, jako jsou listy a stonky.[9] Jako celkový vzorec se délky kořenů snižují s rostoucí koncentrací živin.[4] Vysoké koncentrace minerální živiny, které vedou k toxicita může mít silný vliv také na růst a rozdělení. Například v toxických koncentracích kadmia a olova Fagus sylvatica Bylo prokázáno, že vyvíjí kompaktnější a méně rozvětvující se kořenový systém při vytváření několika kořenových chloupků.[10] Tato změněná struktura funguje tak, že zmenšuje povrch kořenů a účinky toxických živin a zároveň zachovává biomasu pro ty části rostliny, kde by byl růst příznivější.
Voda
Rostliny pěstované v suchých podmínkách mají často sníženou celkovou produkci biomasy, ale také více přispívají ke kořenům a vytvářejí vyšší poměr kořenů a výhonků.[4] Když rostliny přidělují více své biomasy ke kořenům, jsou schopny zvýšit absorpci vody poklepáním dále dolů do vodní stůl a další rozšíření kořenové hmoty příčně. Zvýšené kořenové chloupky také pomáhá při zvyšování absorpce. V případě extrémního sucha v půdě nedochází ke zvýšení kořenů ke střílení biomasy, protože je přijat stav vegetačního klidu.[11] Dostupnost vody také ovlivňuje povrchovou plochu listů, protože příliš velká povrchová plocha by mohla umožnit nadměrnou transpiraci za podmínek s nízkou vodou.
Biotické faktory dělení
Interakce mezi organismy mohou také změnit rozdělení v důsledku konkurence o zdroje, sdílení zdrojů nebo snížení rostlinné biomasy.
Soutěž
Mezidruhové a vnitrodruhové konkurence může způsobit pokles dostupných zdrojů pro jednotlivou rostlinu a změnit způsob rozdělení biomasy. Například konkurence mezi rostlinami způsobuje snížený radiální růst větví a stonku, zatímco zvyšuje růst kořenů a výšky stonku.[12] To poskytuje zdůvodnění důležitosti fenotypu plasticita při udržování kondice v konkurenčním prostředí; čím lépe bude rostlina měnit svoji morfologii, tím bude konkurenceschopnější.
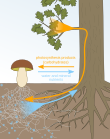
Mutualismus s Mycorrhizae
Vztah mezi rostlinami a mykorhizní houby je příkladem vzájemnost protože rostliny poskytují houbám sacharidy a mykorhizní houby pomáhají rostlinám absorbovat více vody a živin. Vzhledem k tomu, že mykorhizní houby zvyšují absorpci podzemních zdrojů rostlinami, stimulovaly rostliny, které vytvářejí vzájemný vztah s houbami, růst výhonků a vyšší poměr výhonků a kořenů.[13]
Bylinožravý
Bylinožravý způsobí krátkodobé snížení listové hmoty a / nebo stonkové hmoty, které posune stabilitu a proporce částí rostlin. Aby se obnovila rovnováha mezi příjmem živin / vody a rychlostí fotosyntézy, rostliny posílají více energie stonkům a listům. Bylina způsobující vysokou úroveň defoliace (více než 25% plochy listu) zvyšuje růst výhonku a snaží se dosáhnout stejného poměru výhonku a kořene jako před defoliaci.[14] Defoliace proto také omezuje růst kořenů a absorpci živin, dokud se neobnoví poměr kořenů a výhonků před defoliací.
Typ rostliny
Protože různé rostliny mají různé struktury a růstové formy, není ani jejich rozdělení biomasy identické. Například, evergreen stromy mají vyšší hmotnostní frakce listů (LMF) ve srovnání s opadavý stromy. Poměr kmene a kořene se navíc u jednotlivých druhů výrazně liší bylinný rostliny než dřeviny, protože plasticita kořenů v bylinách je vyšší. Dále bylinná jednoděložní rostliny nebo graminoidy ve srovnání s bylinami dvouděložné rostliny mají větší poměr kořenů k celkové hmotnosti (RMF).[15] Rozdíl v bylinných jednoděložných rostlinách a dvouděložných rostlinách lze vysvětlit rozdíly v účinnosti příjmu živin nebo potřebou graminoidů ukládat více škrobu a živin, aby se znovu rozrostly po požáru nebo pastvě. Rozdělení biomasy může být také ovlivněno metodou uhlíkové fixace rostliny: C3 nebo C4. U bylinných jednoděložných rostlin druh fotosyntézy neovlivňuje percentil LMF nebo RMF, ale u bylinných dikot mají druhy C4 nižší RMF.[15] Životní cyklus rostlin může také způsobit různé alokační strategie a poměry listů, kořenů a stonků; ve srovnání se stejnou velikostí eudicotelydonous trvalky, ročníky vložit více energie do rostoucích listů a stonků než u vytrvalých druhů.[15]
Stáří
Stárnoucí vzory biomasy se mohou také měnit, jak rostliny stárnou. v pinus strobus u porostů ve věku 2, 15, 30 a 65 let byl poměr kořene k výhonku 0,32, 0,24, 0,16 a 0,22, což ukazuje pokles kořenové hmoty ve srovnání s hmotou výhonků v prvních několika desetiletích růstu.[16] Věk také ovlivňuje způsob, jakým stromy rozdělují biomasu na různé složky kmene. Růst větví a povrchů listů se s věkem snižuje, jak se dělení na kmen zvyšuje.[16]
Metody / měření
Skládá se z mnoha studií rozdělování biomasy manipulativní experimenty které zvyšují nebo snižují úroveň zdrojů omezujících růst a sledují účinky. Tyto studie často používají rostliny v květináčích pěstovaných ve skleníku k měření účinků hustoty, přebytečných živin, slabého osvětlení atd. Jiné studie se mohou zaměřit na pozorování a analýzu přirozeně se vyskytujících rostlin nebo použít analýzu dat předchozích měření. Metody a měření pro určování dělení biomasy mohou být docela obtížné vzhledem k hmotnosti a objemu větších rostlin. Kromě toho měření velikosti a hmotnosti kořenů vyžaduje promytí a pečlivé odstranění půdy, aby bylo možné dosáhnout přesných měření.[2] Rostlinná biomasa se často měří z hlediska hmotnostního podílu listů (LMF), hmotnostního podílu stonku (SMF) a hmotnostního podílu kořene (RMF), kde je suchá hmotnost rostlinné části nastavena na celkovou suchou hmotnost rostliny.[2] K kvantifikaci rychlosti růstu se používají boční, radiální a výškové přírůstky.
Viz také
Reference
- ^ Farrar, J. F .; Jones, D. L. (2000). "Řízení získávání uhlíku kořeny". Nový fytolog. 147 (1): 43–53. doi:10.1046 / j.1469-8137.2000.00688.x. ISSN 1469-8137.
- ^ A b C d E Poorter, Hendrik; Niklas, Karl J .; Reich, Peter B .; Oleksyn, Jacek; Poot, Pieter; Mommer, Liesje (2012). „Přidělování biomasy listím, stonkům a kořenům: metaanalýzy mezidruhové variace a regulace prostředí“. Nový fytolog. 193 (1): 30–50. doi:10.1111 / j.1469-8137.2011.03952.x. ISSN 1469-8137. PMID 22085245.
- ^ Mohammed, Toungos (06.08.2018). „Látky pro růst rostlin v rostlinné výrobě: recenze“. Citovat deník vyžaduje
| deník =(Pomoc) - ^ A b C Wu, Fuzhong; Bao, Weikai; Li, Fanglan; Wu, Ning (01.05.2008). „Účinky stresu ze sucha a přísun dusíku na růst, rozdělování biomasy a účinnost využívání vody u sazenic Sophora davidii“. Environmentální a experimentální botanika. 63 (1–3): 248–255. doi:10.1016 / j.envexpbot.2007.11.002.
- ^ Ploeg, R. R. van der; Böhm, W .; Kirkham, M. B. (1999). „O původu teorie minerální výživy rostlin a zákonu minima“. Soil Science Society of America Journal. 63 (5): 1055–1062. doi:10,2136 / sssaj1999,6351055x. ISSN 1435-0661.
- ^ Wang, G. Geoff; Bauerle, William L .; Mudder, Bryan T. (2006-05-01). „Účinky aklimatizace světla na fotosyntézu, růst a přidělování biomasy u sazenic kaštanu amerického (Castanea dentata)“. Ekologie a management lesů. 226 (1): 173–180. doi:10.1016 / j.foreco.2005.12.063. ISSN 0378-1127.
- ^ A b Chu, Celia C .; Coleman, James S .; Mooney, Harold A. (04.04.1992). „Kontroly dělení biomasy mezi kořeny a výhonky: Atmosférické obohacení CO2 a získávání a přidělování uhlíku a dusíku v divoké ředkvi“. Ekologie. 89 (4): 580–587. Bibcode:1992Ocol..89..580C. doi:10.1007 / BF00317167. ISSN 1432-1939. PMID 28311891.
- ^ Bernacchi, C. J .; Coleman, J. S .; Bazzaz, F. A .; McConnaughay, K. D. M. (2000). „Přidělování biomasy u jednoletých druhů starých polních rostlin pěstovaných v prostředí se zvýšeným obsahem CO2: žádný důkaz pro optimální rozdělení“. Globální biologie změn. 6 (7): 855–863. Bibcode:2000GCBio ... 6..855B. doi:10.1046 / j.1365-2486.2000.00370.x. ISSN 1365-2486.
- ^ A b Cambui CA; Svennerstam H; Gruffman L; Nordin A; Ganeteg U; et al. (2011). „Vzory rozdělení rostlinné biomasy závisí na zdroji dusíku“. PLOS ONE. 6 (4): e19211. Bibcode:2011PLoSO ... 619211C. doi:10.1371 / journal.pone.0019211. PMC 3081341. PMID 21544211.
- ^ Breckle, S.-W .; Kahle, H. (01.07.1992). „Účinky toxických těžkých kovů (Cd, Pb) na růst a minerální výživu buku (Fagus sylvatica L.)“. Vegetatio. 101 (1): 43–53. doi:10.1007 / BF00031914. ISSN 1573-5052.
- ^ Xu, Zhenzhu; Zhou, Guangsheng; Shimizu, Hideyuki (01.06.2010). „Reakce rostlin na sucho a zavlažování“. Signalizace a chování zařízení. 5 (6): 649–654. doi:10,4161 / psb.5.6.11398. ISSN 1559-2324. PMC 3001553. PMID 20404516.
- ^ Yang, Siao-čou; Zhang, Wen-hui; On, Qiu-yue (01.01.2019). „Účinky vnitrodruhové soutěže na růst, architekturu a přidělování biomasy Quercus Liaotungensis“. Journal of Plant Interactions. 14 (1): 284–294. doi:10.1080/17429145.2019.1629656. ISSN 1742-9145.
- ^ Veresoglou, Stavros D .; Menexes, George; Rillig, Matthias C. (04.04.2012). „Ovlivňují arbuskulární mykorhizní houby alometrické rozdělení biomasy hostitelských rostlin na výhonky a kořeny? Metaanalýza studií od roku 1990 do roku 2010“. Mycorrhiza. 22 (3): 227–235. doi:10.1007 / s00572-011-0398-7. ISSN 1432-1890. PMID 21710352.
- ^ Mihaliak, Charles A .; Lincoln, David E. (01.03.1989). „Rozdělení rostlinné biomasy a chemická obrana: reakce na defoliaci a omezení dusičnanů“. Ekologie. 80 (1): 122–126. Bibcode:1989Oecol..80..122M. doi:10.1007 / BF00789940. ISSN 1432-1939. PMID 23494354.
- ^ A b C Poorter, Hendrik; Jagodzinski, Andrzej M .; Ruiz-Peinado, Ricardo; Kuyah, Shem; Luo, Yunjian; Oleksyn, Jacek; Usoltsev, Vladimir A .; Buckley, Thomas N .; Reich, Peter B .; Sack, Lawren (listopad 2015). „Jak se mění distribuce biomasy s velikostí a liší se mezi druhy? Analýza 1200 druhů rostlin z pěti kontinentů“. Nový fytolog. 208 (3): 736–749. doi:10.1111 / nph.13571. ISSN 0028-646X. PMC 5034769. PMID 26197869.
- ^ A b Peichl, Matthias; Arain, M. Altaf (2007-12-15). „Alometrie a dělení nadzemní a podzemní stromové biomasy ve věkové posloupnosti bílých borovicových lesů“. Ekologie a management lesů. 253 (1): 68–80. doi:10.1016 / j.foreco.2007.07.003. ISSN 0378-1127.