Priming reakce - Response priming
V psychologie vnímání a ovládání motoru, termín primární reakce označuje zvláštní formu priming. Obecně platí, že primingové efekty nastávají vždy, když je reakce na cílový stimul ovlivněna prime podnět předloženo dříve. Charakteristickým rysem aktivace odezvy je to, že primární a cílová jsou prezentovány v rychlém sledu (obvykle méně než 100 milisekund od sebe) a jsou spojeny se stejnými nebo alternativními motorickými odezvami. Když je pro klasifikaci cílového stimulu provedena zrychlená motorická odezva, může prvočíslo bezprostředně předcházející cíli vyvolat konflikty odezvy, pokud je přiřazeno jiné odezvě jako cíl. Tyto konflikty odezvy mají pozorovatelné účinky na chování motoru, což vede k efektům aktivace, např. V dobách odezvy a chybovosti. Zvláštní vlastností odezvy na odezvu je její nezávislost na vizuálním povědomí o náletu.
Priming reakce jako visuomotorický efekt
V roce 1962 Fehrer a Raab[1] hlášeny experimenty, kde se od účastníků vyžadovalo, aby po předložení vizuálního podnětu cíle co nejrychleji stiskli jednu klávesu. Viditelnost cíle byla výrazně snížena tzv. Metakontrastním maskováním (viz níže). Autoři zjistili, že doby odezvy byly nezávislé na subjektivní viditelnosti cíle, tj. Reakce na dobře viditelné cíle byly stejně rychlé jako reakce na téměř neviditelné cíle (Fehrer-Raabův efekt). Termín odezva byla poprvé použita Rosenbaumem a Kornblumem[2] s ohledem na experiment paradigma kde různé aspekty motorických odpovědí byly připraveny vizuálními podněty. Moderní postup reakce primingu vyvinuli v 80. a 90. letech Peter Wolff, Werner Klotz, Ulrich Ansorge a Odmar Neumann na univerzitě v Bielefeldu v Německu.[3][4][5][6] Toto paradigma bylo dále rozvíjeno v 90. letech výzkumným týmem vedeným Dirkem Vorbergem na univerzitě v Braunschweigu v Německu.[7]
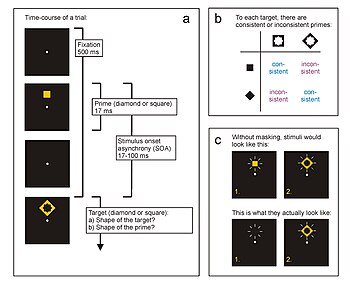
Typický časový průběh pokusu v paradigmatu iniciace odpovědi. Zde účastník reaguje co nejrychleji na tvar cílového stimulu (kosočtverec nebo čtverec) stisknutím přiřazené klávesy odpovědi. Krátce předtím se objeví prvočíslo (také diamant nebo čtverec), které ovlivňuje reakci na cíl. Časový interval mezi hlavním nástupem a nástupem cíle se nazývá „asynchronie nástupu stimulu“ (SOA). V mnoha experimentech s přípravou odpovědi slouží cíl také k vizuálnímu maskování připravenosti. Proto se často používá druhý úkol, kdy jsou účastníci požádáni, aby identifikovali maskované prvočíslo. b) Prime a target jsou konzistentní, pokud jsou přiřazeny ke stejné odpovědi, a nekonzistentní, pokud jsou přiřazeny k různým reakcím. c) Viditelnost prime může být silně ovlivněna maskováním účinků od cíle.

Ve všech paradigmatech přípravy reakce musí účastníci reagovat na konkrétní cílový stimul. V jednoduchém experimentu by to mohl být jeden ze dvou geometrických stimulů, z nichž každý je přiřazen jednomu ze dvou klíčů odezvy (např. Diamant - levý klíč; čtverec - pravý klíč). Experiment se skládá z velkého počtu pokusů, kdy účastník stiskne levou klávesu po objevení diamantu a pravou klávesu po zobrazení čtverce, a to co nejrychleji a nejsprávněji. V každé studii předchází terč prvočíslo, které je také diamantem nebo čtvercem, a proto je schopno vyvolat stejné motorické odezvy jako cíl (obr. 1a). Pokud jsou primární a cílová skupina spojeny se stejnou odpovědí (kosočtverec předchází kosočtverec, čtverec předchází kvadrát), nazývá se „konzistentní“ (také „shodný“, „kompatibilní“); pokud jsou spojeny s různými motorickými odezvami (kosočtverec, kterému předchází čtverec, čtverci, kterému předchází kosočtverec), nazývá se „nekonzistentní“ (také „nesouhlasný“, „nekompatibilní“; obr. 1b). Časový interval mezi nástupem prime a nástupem cíle se nazývá „stimulační nástup asynchronie“ (SOA). Typicky se používají SOA do 100 milisekund (ms).
Primingové efekty nastávají, když prime ovlivňuje motorickou odezvu na cíl: konzistentní připraví rychlostní reakce na cíl, zatímco nekonzistentní připraví pomalé reakce (obr. 2). Účinky nasávání v dobách odezvy se počítají z rozdílu mezi dobami odezvy v nekonzistentních a konzistentních studiích. Kromě jejich účinků na rychlost odezvy mohou prvočísla významně ovlivnit míru chyb odezvy (tj. Chybné reakce na cíl): konzistentní prvočísla zřídka vedou k chybám, zatímco míra chyb může u nekonzistentních prvočísel být velmi vysoká. V dobách odezvy i v četnosti chyb se účinky primingu obvykle zvyšují s SOA, což vede k typickým vzorům aktivace odpovědi na obr.[7] To znamená, že čím více času mezi hlavním a cílovým časem uplyne, tím větší jsou vlivy hlavního proudu na reakci. Při průměrných dobách odezvy 350-450 ms může být efekt priming reakce až 100 ms, což z něj činí jeden z numericky největších efektů ve výzkumu doby odezvy.
Výsledky mnoha experimentů ukazují, že ke zvýšení primingu pomocí SOA dochází, protože prime má zvyšující se čas na ovlivnění procesu odezvy, než může vstoupit do hry skutečný cílový stimul a řídit motorickou odezvu samostatně. To je patrné z časového průběhu motorické aktivity v EEG,[8][9][10][11][12] z nasměrovaných odezvových reakcí,[13][14][15] z měření síly odezvy,[16] a ze simulačních studií,[7] což vše naznačuje, že k aktivaci motoru dochází nejprve ve směru určeném prvočíslem, a teprve poté probíhá ve směru určeném skutečným cílem. Proto konečná velikost efektu primingu závisí jak na vlastnostech stimulů, tak na vlastnostech úkolu. Prvočísla s vysokou stimulační energií (např. Vyšší kontrast, delší trvání) a úkoly se snadnou diskriminací stimulů vedou k velkým efektům nasávání, zatímco prvočísla s nízkou stimulační energií a úkoly s obtížnou diskriminací vedou k menším účinkům.[14][15] Účinky nasávání mohou být zesíleny vizuální pozorností zaměřenou na pozici prime nebo na jeho relevantní rysy právě včas pro vzhled prime.[17][18][19]
Doposud popsaný časový průběh efektu aktivace odezvy platí pouze pro SOA do asi 100 ms. U delších SOA se může primární účinek dále zvýšit. Za určitých okolností však lze pozorovat obrácení účinku, když nekonzistentní prvočísla vedou k rychlejší reakci na cíl než konzistentní prvočísla. Tento efekt se nazývá „negativní účinek kompatibility“.[9][11][20][21][22][23][24]
Maskovaný základ
K vyšetřování jevů vnímání bez lze použít primární reakci povědomí. Zde lze viditelnost prvočísla systematicky snížit nebo dokonce zrušit pomocí maskujícího podnětu. Toho lze dosáhnout předložením maskujícího stimulu krátce před nebo po prvočísle.[25] Viditelnost prime může být posouzena různými opatřeními, jako jsou úkoly diskriminace vynucené volby, rozsudky detekce stimulů, úsudky jasu a další.[26][27]Fenomén jménem metakontrast maskování lze vytvořit, když za prvočíslem následuje maska obklopující tvar prvočísla, takže oba podněty sdílejí sousední kontury. Například prvočíslo může být maskováno větším prstencem, jehož vnitřní obrysy přesně zapadají do tvaru prvočísla. V mnoha experimentech s aktivací odezvy slouží cíl dalšímu účelu maskování prime (obr. 1). Metacontrast je forma vizuálu zpětné maskování kde viditelnost prvočísla je snížena podnětem následujícím po prvočísle.[25][28][29]
Na obr. 3 jsou ukázány některé typické časové průběhy vizuálních maskovacích efektů jako funkce primární cílové SOA v experimentu vyvolávajícím reakci, kde samotný cíl slouží jako maskující stimul (obr. 1a, c). Tady by míra prvotřídní viditelnosti mohla být diskriminace výkon účastníka, který se snaží uhodnout tvar prvočísla (diamant nebo čtverec) v každém pokusu. Bez maskování by byl výkon téměř dokonalý; účastník by měl při každé zkoušce jen malé potíže se správnou klasifikací jako čtverce nebo diamantu. Naproti tomu, pokud by bylo maskování dokončeno, diskriminační výkon by byl na úrovni náhody (obr. 3, levý panel). V mnoha experimentech je však časový průběh maskování méně extrémní (obr. 3, pravý panel). Většina stimulačních podmínek vede k takzvanému „maskování typu A“, kde je stupeň maskování nejvyšší při krátkých SOA a poté se snižuje, takže prime je snazší rozlišovat pro zvyšování SOA. Za určitých okolností však lze získat „maskování typu B“, kde je stupeň maskování nejvyšší u středně pokročilých SOA, ale kde se prime lépe snáze rozlišuje při kratších nebo delších SOA. K maskování typu B může dojít při maskování metakontrastem, ale kriticky závisí na stimulačních vlastnostech prvočísel a cílů.[25][28] Časový průběh maskování se navíc může velmi lišit od člověka k člověku.[30]
Nezávislost reakce od vizuálního vědomí

Experimenty ukazují, že časový průběh účinku primární reakce (zvyšující se účinky se zvyšující se SOA) je nezávislý na stupni a časovém průběhu maskování. Klotz a Neumann (1999) demonstrovali účinky primingové odpovědi pod úplným maskováním prime.[4] Rozšíření těchto zjištění na další časové průběhy vizuálního maskování, Vorberg et al.[7] měnil časový průběh maskování řízením relativního trvání prvočísel a cílů. Ve svých experimentech byly cíli šipky směřující doleva nebo doprava, zatímco prvočísla byly menší šipky metakontrastně maskované cíli. Když se účastníci pokusili identifikovat směr ukazování prvočísel, bylo možné vytvořit jakýkoli z časových průběhů zobrazených na obr. 3 v závislosti na podmínkách stimulu: úplná viditelnost, úplné maskování, maskování typu A a maskování typu B. Když však účastníci museli co nejrychleji reagovat na směr cíle, byly primovací účinky za všech těchto podmínek prakticky totožné. Kromě toho byl časový průběh primingu vždy stejný (zvyšující se efekt primingu se zvyšujícím se SOA), bez ohledu na to, zda byla prvočísla viditelná nebo neviditelná, a bez ohledu na to, zda se viditelnost zvýšila nebo snížila pomocí SOA.[7]
Zvláštní pozornost je třeba věnovat případům, kdy se účinky primingu zvyšují, i když se snižuje viditelnost. Takový opačný časový průběh nasávání a vizuální uvědomování si prvočísel jasně ukazují, že oba procesy spoléhají na odlišné mechanismy.[31] Toto zjištění by mohlo být potvrzeno v mnoha dalších experimentech, které odhalily četné disociace mezi maskováním a primováním.[1][3][4][5][13][14][19][32][33][34][35][36][37] Nezávislost nasávání a vizuálního povědomí jasně odporuje tradiční představě, že účinky nevědomého vnímání pouze odrážejí určitou zbytkovou schopnost zpracování za velmi nepříznivých podmínek sledování, něco, co přetrvává i poté, co byl primární podnět degradován tak silně, že jeho vědomé povědomí pokleslo pod některé "práh". Tato koncepce často vedla k ostré kritice výzkumu nevědomí nebo „podprahové“ vnímání,[38][39][40][41] ale na základní úrovni je to pravděpodobně špatně. Místo toho je motorická aktivace maskovanými prvočísly zjevně nezávislá na procesech zpětného maskování za předpokladu, že viditelnost je řízena pouze maskujícím stimulem, zatímco primární stimul zůstává nezměněn. Jinými slovy: na krátkou dobu a za vhodných experimentálních podmínek mohou vizuálně maskované (neviditelné) podněty ovlivňovat motorické reakce stejně účinně jako viditelné.
Varianty
Vzhledem k tomu, že si výzkumník je vědom nejvlivnějších experimentálních proměnných,[42] metoda iniciace reakce může být použita v řadě experimentálních variant a může přispět k prozkoumání mnoha výzkumných otázek v oblasti kognitivní psychologie.[43]Nejběžnější forma odezvy na primární reakci využívá primární a cílový bod na stejné pozici monitoru, takže cíl slouží také k vizuálnímu maskování primárního bodu (často pomocí metakontrastu). V mnoha experimentech existují dva různé cíle, kterým předcházejí dvě různá prvočísla na stejných pozicích monitoru.[3][4][13] Účastníci pak musí rozlišit dva cíle a reagovat na pozici cíle relevantního pro daný úkol. Někdy se používají tři typy stimulů (prime, mask, target), zvláště když musí být SOA prime-target velmi dlouhá.[21][22] Někdy není použita vůbec žádná maska.[19] Prvočísla a cíle se nemusí zobrazovat na stejné pozici obrazovky: Jeden stimul může sousedit s druhým, jako v Eriksenovo paradigma[7][11][44][45] (ve skutečnosti může být Eriksenův efekt zvláštním případem primování odpovědi).
Účinky vyvolávající reakci byly prokázány u velkého počtu podnětů a diskriminačních úkolů, včetně geometrických podnětů,[3][4][15] barevné podněty,[13][14][32] různé typy šípů,[7][9][10][46] přírodní obrazy (zvířata vs. objekty),[15] samohlásky a souhlásky,[47] písmena,[44] a číslice.[33] V jedné studii byly šachové konfigurace prezentovány jako prvočísla a cíle a účastníci se museli rozhodnout, zda je král pod kontrolou.[35] Mattler (2003) by mohl ukázat, že odezva priming může nejen ovlivnit motorické odezvy, ale funguje také pro kognitivní operace, jako je prostorový posun vizuální pozornosti nebo posun mezi dvěma různými úkoly doby odezvy.[36] Byly použity také různé typy maskování. Namísto měření klávesových odpovědí (obvykle se dvěma alternativami odezvy) používají některé studie více než dvě alternativy odezvy nebo zaznamenávají odezvy na řeč,[5] zrychlené pohyby prstem,[13][14][15] pohyby očí,[48] nebo takzvané potenciály připravenosti, které odrážejí stupeň motorické aktivace v motorické kůře mozku a lze je měřit elektro-encefalografické metody.[8][9][12][49] Metody zobrazování mozku jako funkční zobrazování magnetickou rezonancí (fMRI) byly také použity.[33]
Teorie
V části, která následuje, budou popsány tři teorie, které vysvětlují pravidelné, primární reakce na pozitivní reakci. Přehled teorií o negativním vlivu kompatibility lze nalézt v Sumner (2007).[24]
Přímá specifikace parametrů

Teorii přímé specifikace parametrů (obr. 4) navrhl Odmar Neumann z univerzity v Bielefeldu v Německu, aby vysvětlil Fehrer-Raabův efekt a také výsledky studií časné odezvy.[50] Teorie předpokládá, že na začátku experimentu s primingem odpovědi účastníci získají pravidla přiřazení stimul-odpověď, která se rychle automatizují. Po této praktické fázi lze motorickou odezvu připravit tak daleko, že ke specifikaci odezvy je stále zapotřebí pouze jeden kritický stimulační prvek (např. Diamant vs. čtverec). Tato funkce příchozího stimulu poté definuje poslední chybějící parametr akce (např. Stisknutí klávesy vlevo vs. vpravo). Odpovědi jsou vyvolávány rychle a přímo, bez nutnosti vědomé reprezentace vyvolávajícího podnětu. Priming odezvy je vysvětlen za předpokladu, že vlastnosti prime vyvolávají přesně stejné procesy specifikace parametrů, které mají být vyvolány cílovým stimulem. Souběžně s procesem vyvolávání odezvy se objevuje vědomé znázornění prvočísel a cílů, které lze podrobit vizuálním maskovacím procesům. Vědomá reprezentace stimulů však v současné experimentální studii nehraje pro motorické procesy žádnou roli.
Účet spouštěče akce
Účet spouštěče akce byl vyvinut společností Wilfried Kunde, Andrea Kiesel a Joachim Hoffmann na univerzitě ve Würzburgu v Německu.[47][51] Tento účet předpokládá, že odpovědi na bezvědomá prvočísla nejsou ani vyvolány sémantický analýza prvočísel ani předem stanovená mapování stimul-odezva. Místo toho se předpokládá, že prime odpovídá podmínce vydání již existující akce a vyvolá přiřazenou odpověď jako klíč otevírající zámek. To se děje ve dvou po sobě jdoucích krocích. V prvním kroku jsou spouštěče akcí drženy aktivní v pracovní paměti, které odpovídají příslušnému úkolu a jsou schopné vyvolat konkrétní motorickou odezvu. Spouštěče akcí jsou stanoveny ve fázi výuky a cvičení experimentu. Ve druhém kroku zvaném online zpracování stimulů je nadcházející stimul srovnáván s podmínkami uvolnění akce. Pokud stimul vyhovuje podmínkám spouštění, spouštění akce automaticky provede odpověď. Úkolem účastníka může být například označení, zda je vizuálně prezentované číslo menší nebo větší než pět,[33] tak, že je stisknuto levé tlačítko pro čísla „1“ až „4“ a pravé tlačítko pro čísla „6“ až „9“. Na základě instrukce jsou nastaveny spouštěče akcí, které automaticky vyvolávají odpověď přiřazenou prvočíslu nebo cílovému číslu. Jednou důležitou předpovědí této teorie je, že reakce mohou být vyvolány prvočísly, která splňují spouštěcí podmínky, ale nikdy se neobjeví jako cíle.[51]
Vědomá reprezentace stimulu opět nehraje žádnou roli pro motorickou aktivaci; Může to však vést ke strategické úpravě kritérií odezvy v pozdějších studiích (např. volbou pomalejší reakce, aby nedocházelo k chybám). V souhrnu lze tuto teorii považovat za rozšiřující koncept přímé specifikace parametrů zaměřením na přesné podmínky, které vedou k priming odpovědi.
Teorie rychlého pronásledování
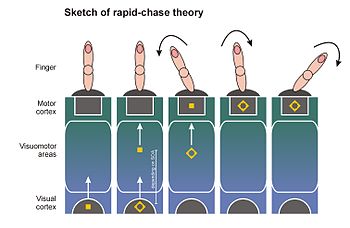
Teorie rychlého pronásledování primární reakce[12][14][15] v roce 2006 navrhli Thomas Schmidt, Silja Niehaus a Annabel Nagel. Vazuje model přímé specifikace parametrů na zjištění, že nově se vyskytující vizuální podněty vyvolávají vlnu aktivace neuronů ve visuomotorickém systému, která se rychle šíří z vizuální na motorické oblasti kůry.[52][53][54][55] Protože vlnoplocha aktivity se šíří velmi rychle, navrhli Victor Lamme a Pieter Roelfsema z Amsterdamské univerzity, aby tato vlna začala jako čistý proces dopředné reakce (dopředné zametání ): Buňka, kterou nejprve dosáhne vlnoplocha, musí předat svoji aktivitu, než bude schopna integrovat zpětnou vazbu z jiných buněk. Lamme a Roelfsema předpokládají, že tento druh dopředného zpracování není dostatečný k vytvoření vizuálního povědomí o stimulu: neuronální zpětná vazba a jsou vyžadovány smyčky pro opakované zpracování, které spojují rozšířené neuronové sítě.[29][53]
Podle teorie rychlého pronásledování jak prvočísla, tak i cíle vyvolávají dopředné zatáčky, které procházejí visuomotorickým systémem v rychlém sledu, dokud nedosáhnou motorických oblastí mozku. Tam jsou motorické procesy vyvolávány automaticky a bez nutnosti vědomé reprezentace. Vzhledem k tomu, že primární signál má náskok nad cílovým signálem, jsou prvočísla a cíle zapojeny do „rychlého pronásledování“ prostřednictvím visuomotorického systému. Protože primární signál dosáhne motorické kůry jako první, je schopen aktivovat přiřazenou motorickou odezvu. Čím kratší je SOA hlavního cíle, tím rychleji může cíl zahájit pronásledování. Když cílový signál konečně dorazí do motorické kůry, může pokračovat v procesu odezvy vyvolaném prvočíslem (pokud je prvočíslo a cíl konzistentní) nebo přesměrovat proces odezvy (pokud jsou prvočíslo a cíl nekonzistentní). To vysvětluje, proč se efekty primingu odezvy zvyšují s SOA prime-target: čím delší je SOA, tím více času pro primární signál k řízení reakce samostatně, a dále může proces aktivace odezvy pokračovat ve směru prime. Za určitých okolností může prvočíslo také vyvolat chybu odezvy (vedoucí k charakteristickým účinkům primování v chybovosti). Takový časový průběh sekvenčního řízení motoru prvočísly a cíli popsal v roce 2003 Dirk Vorberg a spolupracovníci v matematickém modelu[7] a odpovídá časovému průběhu aktivovaných motorických potenciálů v EEG.[8][12][56]
Podle teorie rychlého pronásledování jsou účinky iniciace reakce nezávislé na vizuálním povědomí, protože jsou přenášeny procesy rychlé dopředné reakce, zatímco vznik vědomé reprezentace stimulů závisí na pomalejších, opakujících se procesech.[29][53]Nejdůležitější predikcí teorie rychlého pronásledování je, že dopředné tahy hlavních a cílových signálů by měly probíhat v přísném pořadí. Tato přísná posloupnost by měla být pozorovatelná v časovém průběhu motorické odezvy a měla by existovat raná fáze, kdy je odezva řízena výhradně prvočíslem a je nezávislá na všech vlastnostech skutečného cílového stimulu. Jedním ze způsobů, jak zkontrolovat tyto předpovědi, je prozkoumat časový průběh odezvových polohovacích odpovědí.[13] Ukázalo se, že tyto odezvové reakce začínají ve stanoveném čase po prezentaci prime (nikoli skutečného cíle) a začínají postupovat ve směru určeném prime. Pokud je hlavní a cílový objekt nekonzistentní, je cíl často schopen obrátit směr ukazování „za běhu“ a směřovat reakci správným směrem. Čím delší je však SOA, tím delší je čas, kdy se prst pohybuje ve směru zavádějícího prime.[13] Schmidt, Niehaus a Nagel (2006) by mohli ukázat, že nejranější fáze nasměrovaných pohybů zaměřování výhradně závisí na vlastnostech prime (např. Barevný kontrast červené vs. zelené prvočísla), ale je nezávislá na všech vlastnostech cíle ( čas jeho výskytu, jeho barevný kontrast a schopnost maskovat prvočíslo).[14] Tyto nálezy lze potvrdit různými metodami a různými typy podnětů.[12][15][18][19]
Vzhledem k tomu, že teorie rychlého pronásledování pohlíží na primární odezvu jako na proces dopředné reakce, tvrdí, že k aktivačním účinkům dochází dříve, než se na zpracování stimulu zúčastní opakující se a zpětnovazební aktivita. Tato teorie proto vede ke kontroverzní tezi, že účinky primingů odezvy jsou mírou předvědomého zpracování vizuálních podnětů, které se mohou kvalitativně lišit od způsobu, jakým jsou tyto podněty konečně reprezentovány ve vizuálním vědomí.[37]
Viz také
- Precizní zpracování
- Teorie nevědomého myšlení # Kritika UTT
- Vizuální maskování
- Sluchové maskování
- Pozorné mrknutí
Reference
- ^ A b Fehrer, E. a Raab, D. Reakční doba na podněty maskované metakontrastem. V: Journal of Experimental Psychology, Nr. 63, 1962, str. 143-147.
- ^ Rosenbaum, D. A., & Kornblum, S. (1982). Primovací metoda pro vyšetřování výběru motorických odpovědí. Acta Psychologica, 51, str. 223-243.
- ^ A b C d Klotz, W., & Wolff, P.: Účinek maskovaného stimulu na reakci na maskující stimul. V: Psychologický výzkum, Nr. 58, 1995, s. 92-101.
- ^ A b C d E Klotz, W. a Neumann, O.: Aktivace motoru bez vědomé diskriminace při maskování metakontrastu. V: Journal of Experimental Psychology: Human Perception and Performance, Nr. 25, 1999, s. 976-992.
- ^ A b C Ansorge, U., Klotz, W., &, O.: Ruční a slovní reakce na zcela maskované (nehlášené) podněty: Zkoumání některých podmínek pro disociaci metakontrastu. V: Vnímání, Nr. 27, 1998, s. 1177-1189.
- ^ Ansorge, U., Neumann, O., Becker, S.I., Kälberer, H., & Kruse, H.: Senzomotorická nadvláda: Vyšetřování vědomého a nevědomého vidění maskovaným primováním. V: Pokroky v kognitivní psychologii, Nr. 3, 2007, s. 257-274.
- ^ A b C d E F G h Vorberg, D., Mattler, U., Heinecke, A., Schmidt, T., & Schwarzbach, J .: Různé časové kurzy vizuálního vnímání a akčního primingu. V: Sborník Národní akademie věd USA, Nr. 100, 2003, s. 6275-6280.
- ^ A b C Leuthold, H., & Kopp, B.: Mechanismy aktivace maskovanými podněty: Odkazy z mozkových potenciálů souvisejících s událostmi. V: Psychologická věda, Nr. 9, 1998, s. 263-269.
- ^ A b C d Eimer, M., & Schlaghecken, F.: Účinky maskovaných stimulů na motorickou aktivaci: Behaviorální a elektrofyziologické důkazy. V: Journal of Experimental Psychology: Human Perception and Performance, Nr. 24, 1998, s. 1737-1745.
- ^ A b Eimer, M., & Schlaghecken, F.:Usnadnění reakce a inhibice v podprahové aktivaci. V: Biologická psychologie, Nr. 64, 2003, s. 7-26.
- ^ A b C Mattler, U .: Zpožděné účinky flankerů na potenciály laterální připravenosti. V: Experimentální výzkum mozku, Nr. 151, 2003, str. 272-288.
- ^ A b C d E Vath, N., & Schmidt, T.: Sledování sekvenčních vln rychlé visuomotorické aktivace v potenciálech laterální připravenosti. V: Neurovědy, Nr. 145, 2007, s. 197-208.
- ^ A b C d E F G Schmidt, T .: Prst v letu: Ovládání motoru v reálném čase vizuálně maskovanými barevnými podněty. V: Psychologická věda, Nr. 13, 2002, S. 112-118.
- ^ A b C d E F G h Schmidt, T., Niehaus, S., & Nagel, A .: Prvočísla a cíle v rychlých honičkách: Sledování postupných vln aktivace motoru. V: Behaviorální neurovědy, Nr. 120, 2006, s. 1005-1016.
- ^ A b C d E F G Schmidt, T., & Schmidt, F .: Zpracování přirozených obrázků je dopředné: Jednoduchý test chování. V: Pozornost, vnímání a psychofyzika, Nr. 71, 2009, s. 594-606.
- ^ Mattler, U .: Flankerovy účinky na výkon motoru a hypotéza aktivace reakce na pozdní úrovni. V: Quarterly Journal of Experimental Psychology, Nr. 58A, 2005, s. 577-601.
- ^ Sumner, P., Tsai, P.-C., Yu, K., & Nachev, P .: Pozorná modulace senzomotorických procesů při absenci vnímavého vědomí. V: Sborník Národní akademie věd USA, Nr. 103, 2006, s. 10520-10525.
- ^ A b Schmidt, T., & Seydell, A .: Vizuální pozornost zesiluje aktivaci odezvy zaměřovacích pohybů na barevné cíle. V: Vnímání a psychofyzika, Nr. 70, 2008, s. 443-455.
- ^ A b C d Schmidt, F., & Schmidt, T .: Zvláštní pozornost věnovaná tvarům a barvám v bezvědomí. V: Pozornost, vnímání a psychofyzika, Nr. 72, 2010, s. 1480-1494.
- ^ Jaskowski, P .: Negativní účinek kompatibility s nemaskujícími se hranami: Případ hypotézy inhibice vyvolané maskou. V: Vědomí a poznání, Nr. 17, 2008, s. 765-777.
- ^ A b Verleger, R., Jaskowski, P., Aydemir, A., van der Lubbe, R. H. J., & Groen, M .: Kvalitativní rozdíly mezi vědomým a nevědomým zpracováním? Při inverzní aktivaci vyvolané maskovanými šipkami. V: Journal of Experimental Psychology, Nr. 133, 2004, s. 494-515.
- ^ A b Lingnau, A., & Vorberg, D .: Časový průběh inhibice odezvy v maskovaném primingu. V: Vnímání a psychofyzika, Nr. 67, 2005, s. 545-557.
- ^ Klapp, S. T., & Hinkley, L. B .: Negativní účinek kompatibility: Inhibice v bezvědomí ovlivňuje reakční dobu a výběr odezvy. V: Journal of Experimental Psychology: General, Nr. 131, 2002, s. 255-269.
- ^ A b Sumner, P .: Negativní a pozitivní maskování - důsledky pro motorickou inhibici. V: Pokroky v kognitivní psychologii, Nr. 3, 2007, s. 317-326.
- ^ A b C Breitmeyer, B. G., & Öğmen, H .: Vizuální maskování. V: Scholarpedia, 2007, 2(7): 3330.
- ^ Ansorge, U., Breitmeyer, B. G., & Becker, S. I .: Porovnání citlivosti napříč různými měrami zpracování za podmínek maskování metakontrastu. V: Vision Vision, Nr. 47, 2007, s. 3335-3349.
- ^ Ansorge, U., Becker, S. I., & Breitmeyer, B .:Přehodnocení disociace metakontrastu: Porovnání citlivosti mezi různými opatřeními a úkoly. V: Quarterly Journal of Experimental Psychology, Nr. 62, 2009, s. 286-309.
- ^ A b Breitmeyer, B. G., & Öğmen, H .: Vizuální maskování. Oxford: Oxford University Press, 2006
- ^ A b C DiLollo, V., Enns, J. T., & Rensink, R. A .: Soutěž o vědomí mezi vizuálními událostmi: Psychofyzika nových vizuálních procesů. V: Journal of Experimental Psychology: General, Nr. 129, 2000, s. 481-507.
- ^ Albrecht, T., Klapötke, S., & Mattler, U .: Individuální rozdíly v maskování metakontrastu jsou vylepšeny percepčním učením. V: Vědomí a poznání, Nr. 19. 2010, s. 656–666.
- ^ Schmidt, T., & Vorberg, D .: Kritéria pro nevědomí: Tři typy disociace. V: Vnímání a psychofyzika, Nr. 68, 2006, s. 489-504.
- ^ A b Breitmeyer, B. G., Ro, T., & Singhal, N. S .: Nevědomá barevná aktivace nastává na úrovni vizuálního zpracování, která není závislá na vnímání. V: Psychologická věda, Nr. 15, s. 198-202.
- ^ A b C d Dehaene, S., Naccache, L., Le Clec'H, G., Koechlin, E., Mueller, M., Dehaene-Lambertz, G., van de Moortele, P. F. a Le Bihan, D .: Zobrazování sémantické primingu v bezvědomí. V: Příroda, Nr. 395, 1998, s. 597-600.
- ^ Fellows, S., Tabaza, R., Heumann, M., Klotz, W., Neumann, O., Schwarz, M., Noth, J., & Topper, R .: Modifikace funkčního motorického úkolu nevědomě vnímanými smyslovými podněty. V: NeuroReport, Nr. 13, 2002, s. 637–640.
- ^ A b Kiesel, A., W., Pohl, C., Berner, M. P., & Hoffmann, J .: Podvědomě hrající šachy. V: Journal of Experimental Psychology: Learning, Memory, and Cognition, Nr. 35, 2009, s. 292-298.
- ^ A b Mattler, U .: Aktivace mentálních operací maskovanými podněty. V: Vnímání a psychofyzika, Nr. 65, 2003, s. 167-187.
- ^ A b Schmidt, T., Miksch, S., Bulganin, L., Jäger, F., Lossin, F., Jochum, J., & Kohl, P .: Priming reakce poháněný místním kontrastem, nikoli subjektivním jasem. V: Pozornost, vnímání a psychofyzika, Nr. 72, 2010, s. 1556-1568.
- ^ Eriksen, C. W .: Diskriminace a učení bez povědomí: Metodický průzkum a hodnocení. V: Psychologický přehled, Nr. 67, 1960, str. 279-300.
- ^ Holender, D .: Sémantická aktivace bez vědomé identifikace v dichotickém poslechu, parafoveálním vidění a vizuálním maskování: Průzkum a hodnocení. V: Behaviorální a mozkové vědy, Nr. 9, 1986, s. 1-23.
- ^ Holender, D., & Duscherer, K .: Nevědomé vnímání: Potřeba změny paradigmatu. V: Vnímání a psychofyzika, Nr. 66, 2004, s. 872-881.
- ^ Cardoso-Leite, P., & Gorea, A .: K percepční / motorické disociaci: Přehled konceptů, teorie, experimentálních paradigmat a interpretací dat. V: Vidět a vnímat, Nr. 23, 2010, s. 1. 89-151.
- ^ Schmidt, F., Haberkamp, A., & Schmidt, T .: Dávky a nedělat při výzkumu priming reakce. V: Pokroky v kognitivní psychologii, Nr. 7, 2011, s. 120–131.
- ^ Schmidt, T., Haberkamp, A., Veltkamp, G. M., Weber, A., Seydell-Greenwald, A., & Schmidt, F .: Vizuální zpracování v systémech rychlého pronásledování: zpracování obrazu, pozornost a povědomí. V: Hranice v psychologii, Nr. 2, 2011, S. 1–16.
- ^ A b Eriksen, B. A. a Eriksen, C. W .: Účinky šumových písmen na identifikaci cílového písmene v úloze, která není předmětem hledání. V: Vnímání a psychofyzika, Nr. 16, 1974, s. 143-149.
- ^ Schwarz, W., & Mecklinger, A .: Vztah mezi identifikovatelností flanker a účinkem kompatibility. V: Vnímání a psychofyzika, Nr. 57, 1995, s. 1045-1052.
- ^ Jaskowski, P., & Slosarek, M .: Jak důležitý je gestalt prime pro podprahové naplnění? V: Vědomí a poznání, Nr. 16. 2007, s. 485,497.
- ^ A b C d Kunde, W., Kiesel, A., Hoffmann, J .: Vědomá kontrola nad obsahem nevědomého poznání. V: Poznání, Nr. 88, 2003, s. 223-242.
- ^ Schwarzbach, J., & Vorberg, D .: Plnění odpovědi s vědomím i bez něj. V: H. Öğmen & B. G. Breitmeyer (Eds.), První polovina sekundy: Mikrogeneze a časová dynamika vizuálních procesů v bezvědomí a při vědomí. Cambridge, MA: MIT Press.
- ^ Jaskowski, P., Bialunska, A., Tomanek, M., & Verleger, R .: Inhibiční procesy vyvolané maskou a distraktorem při aktivaci motorických odpovědí: Studie EEG., V: Psychofyziologie, Nr. 45, 2008, s. 70-85.
- ^ A b Neumann, O .: Přímá specifikace parametrů a koncept vnímání. V: Psychologický výzkum, Nr. 52, 1990, str. 207-215.
- ^ A b C d Kiesel, A., Kunde, W., & Hoffmann, J .: Mechanismy přípravy podprahové reakce. V: Pokroky v kognitivní psychologii, Nr. 1–2, 2007, s. 1 307-315.
- ^ Bullier, J .: Integrovaný model vizuálního zpracování. V: Recenze výzkumu mozku, Nr. 36, 2001, p. 96-107.
- ^ A b C Lamme, V. A. F., & Roelfsema, P. R.: The distinct modes of vision offered by feedforward and recurrent processing. V: Trendy v neurovědách, Nr. 23, 2000, p. 571-579.
- ^ Thorpe, S. J., Fize, D., & Marlot, C.: Speed of processing in the human visual system. V: Příroda, Nr. 381, 1996, p. 520-522.
- ^ VanRullen, R. & Thorpe, S. J.: Surfing a spike wave down the ventral stream. V: Vision Vision, Nr. 42, s. 2593-2615.
- ^ Klotz, W., Heumann, M., Ansorge, U., & Neumann, O. : Electrophysiological activation by masked primes: Independence of prime-related and target-related activities. In: Advances in Cognitive Psychology. Č. 3, 2007, s. 449–465.