RBM10 - RBM10






Motiv 10 vázající RNA je protein, který je kódován RBM10 gen.[5][6][7][8] Tento gen mapuje na X chromozomu v Xp11.23 u lidí. RBM10 je regulátor alternativního sestřihu.[9][10][11] Alternativní sestřih je proces spojený s genovou expresí k produkci více proteinových izoforem z jednoho genu, čímž se vytvoří funkční rozmanitost a buněčná složitost.[12] RBM10 ovlivňuje expresi mnoha genů,[9][10][13][14][15] účast na různých buněčných procesech a drahách, jako je buněčná proliferace a apoptóza.[10][16] Jeho mutace jsou spojeny s různými lidskými chorobami[17][18][19][20][21][22] jako je TARP syndrom,[22][17] vrozená porucha vázaná na X u mužů vedoucí k pre- nebo postnatální letalitě a různé druhy rakoviny u dospělých.[18][19]
Geny a bílkoviny
Gen RBM10 zabírá ~ 41,6 kb a obsahuje 24 exonů. Tento gen je podroben X-inaktivaci,[6][7] ve kterém je jeden ze dvou genů RBM10 v ženských buňkách transkripčně umlčen tvorbou heterochromatinu.
RBM proteiny tvoří velkou rodinu proteinů vázajících RNA (RBP). Existuje 52 RBM proteinů (HGNC: Výbor pro genovou nomenklaturu HUGO), z nichž každý obsahuje jednu až několik domén vázajících RNA nazývaných rozpoznávací motivy RNA (RRM). RBM10 obsahuje dvě RRM (RRM1 a RRM2) a další domény, jako jsou dva zinkové prsty (ZnF), oktamerová repetice (OCRE), tři signály nukleární lokalizace (NLS) a doména bohatá na glycin (G-patch). Aminokyselinová (aa) sekvence RBM10 je mezi savci zachována. Lidská izoforma 1 RBM10 sdílí 96% a 97% sekvenční homologii s sekvencemi myší a potkanů, což naznačuje, že molekulární funkce RBM10 jsou v podstatě stejné u lidí i hlodavců.
RBM10 má více izoforem generovaných prostřednictvím alternativních událostí sestřihu primárního přepisu RBM10. Hlavní izoformy, 1–4, mohou obsahovat sekvenci exonu 4 (77 zbytků) a / nebo zbytek Val odpovídající poslednímu kodonu exonu 10. Isoforma 1 (930 zbytků) obsahuje sekvenci exonu 4 i V354, zatímco izoforma 4 (929 zbytků) neobsahuje tento valinový zbytek. Podobně exon 4 – minus isoforma 3 (853 zbytků) obsahuje V277, zatímco izoforma 2 (852 zbytků) nikoli. Isoforma 5 (995 zbytků) má delší 65-aa N-konec ve srovnání s izoformou 1. Kromě toho automatická výpočetní analýza pomocí nástroje pro predikci genů Gnomon (gen NCBI) ukázala, že může existovat více než 10 různých RBM izoformy.
Funkce
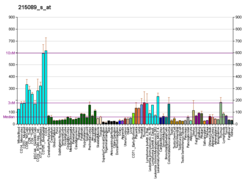
RBM10 je všudypřítomně exprimován téměř v každém typu buněk, jak rostoucích, tak i klidových (UniProtKB-P98175 [člověk] a Q99KG3 [myš]; Atlas lidských proteinů). Obecně je silněji exprimován v aktivních přepisech buněk.[23]
V alternativní regulaci sestřihu podporuje RBM10 vyloučení exonu, nazývaného kazeta nebo alternativní exon, z cílových pre-mRNA a méně často další alternativní sestřihové události, jako je alternativní výběr místa 5ʹ sestřihu.[9][10][11][24] V procesu přeskakování exonů se RBM10 váže blízko k 3lice- a 5ʹ-místům sestřihu kazetových exonů a interferuje s rozpoznáváním a / nebo párováním míst sestřihu, čímž zlepšuje párování míst sestřihu distálně od exonů kazety, což v konečném důsledku vede k vyloučení exonů společně s lemujícími upstream a downstream introny.[9][10][24]
Rozmanitost cílových RNA vázaných RBM10 v buňkách naznačuje, že se účastní různých metabolických procesů, jako je oxidační fosforylace; dráhy spojené s buněčnou proliferací, apoptózou, buněčnou adhezí a reorganizací aktinu / cytoskeletu; a různá onemocnění, jako jsou rakoviny a neurodegenerativní choroby.[10][16][25] Tato data spolu se všudypřítomnou expresí RBM10 naznačují, že se jedná o základní buněčnou složku účastnící se různých buněčných procesů. Kromě alternativní regulace sestřihu se RBM10 účastní i dalších reakcí. Některé příklady jsou polyadenylace srdečních pre-mRNA anti-hypertrofických regulátorů, kde působí jako koregulátor STAR-poly (A) polymerázy,[26] stabilizace mRNA receptoru pro angiotenzin II vazbou na jeho 3ʹ-UTR,[27] biogeneze let-7g miRNA interakcí s jejím předchůdcem,[28] stabilizace p53 vazbou na jeho negativní regulátor, MDM2,[29] zástava buněčného cyklu,[30][31] a antivirové reakce.[32]
RBM10 se lokalizuje v nukleoplazmě, kde dochází k transkripci a sestřihu, stejně jako v membránových jaderných kompartmentech nazývaných jaderná těla S1-1 (S1-1 NB).[23] Počet (přibližně 10–40 na jádro) a velikosti (přibližně 0,5 µm) S1-1 NB se liší podle typu buňky a buněčných podmínek. Když transkripce RNA polymerázy II klesá, RBM10 v nukleoplazmě je izolován v S1-1 NB, které se zvětšují a jsou sférické; po obnovení transkripce se RBM10 a NB S1-1 vrátí do svých počátečních stavů.[23] S1-1 NB se často překrývají s jadernými skvrnami (známé také jako spojovací skvrny nebo shluky interchromatinových granulí),[23][33] zdánlivě naznačuje blízký funkční vztah mezi těmito jadernými doménami, tj. Alternativní regulace sestřihu a sestřihová reakce.
Nařízení
U žen je většina genů na jednom ze dvou chromozomů X transkripčně umlčena tvorbou heterochromatinu a RBM10 je podroben této X-inaktivaci.[6][7][34] Kromě toho existují mechanismy pro řízení zvýšených buněčných hladin RBM10. RBM10 autoreguluje svou nadměrně exprimovanou pre-mRNA alternativním sestřihem, aby se vyloučil exon 6 nebo 12, který generuje předčasný stop kodon v transkriptech, což vede k jejich degradaci nesmyslně zprostředkovaným rozpadem mRNA (NMD).[14] Když transkripce RNA polymerázy II klesá, RBM10 je izolován v S1-1 NB, dokud není transkripce obnovena.[23] Kromě toho RBM10 prochází posttranslačními modifikacemi: fosforylací na mnoha místech v reakci na různé podněty a změny buněčných podmínek (UniProtKB-P98175; PhosphoSitePlus RBM10), stejně jako všudypřítomnost,[35][36] acetylace,[37] a methylace.[38] Molekulární a biologický význam těchto různých posttranslačních modifikací RBM10 však není dobře znám.
Klinický význam
Mutace v RBM10 jsou spojeny s různými lidskými chorobami. Fenotypy způsobené mutacemi RBM10 se liší podle stadií vývoje a postižených tkání. Typickými příklady jsou syndrom TARP, X-vázaná pleiotropní vývojová malformace u novorozenců,[17][22] a různé druhy rakoviny, jako je plicní adenokarcinom (LUAD)[18] a karcinom močového měchýře (BLCA) u dospělých.[19] Tato onemocnění jsou častější u mužů než u žen.[39][40][41] Jedním z důvodů je rozdíl v počtu kopií genu RBM10 v buňce (jedna v mužských buňkách a dvě v ženských buňkách). Mutace v RBM10 se vyskytují v celé molekule a mnoho z nich je nulových mutací. Syndrom TARP je obecně pre- nebo postnatálně smrtelný.[17][42][43] Bylo však hlášeno, že u pacientů ve věku 11, 14 a 28 let tyto nulové mutace unikly.[44][9][45] Mutace RBM10 byly také identifikovány u jiných druhů rakoviny[46] jako jsou renální karcinomy,[47][48][49] rakoviny pankreatu,[50][51] kolorektální rakoviny,[52][53] rakoviny štítné žlázy,[54][55][56] rakoviny prsu,[57] rakoviny žlučovodů,[58][59] rakovina prostaty,[57] a meningiomy nádorů na mozku a astroblastomy.[60][61]
NUMB je nejvíce studovaný downstream efektor RBM10. RBM10 podporuje přeskočení exonu 9 transkriptu NUMB a produkuje izoformu NUMB, která způsobí ubikvitinaci následovanou proteasomální degradací Notch receptoru, a tím inhibuje signální dráhu buněčné proliferace Notch.[10][62][20] U různých druhů rakoviny zvyšují mutace RBM10, které inaktivují nebo snižují jeho alternativní regulační aktivitu sestřihu, produkci exonu 9 - včetně izoformy NUMB, která podporuje proliferaci rakovinných buněk cestou Notch.[10][63][64]
RBM10 potlačuje buněčnou proliferaci[10][27][63][64][65][66][29] a podporuje apoptózu.[27][64][65][29][67][68] Z tohoto důvodu je obecně považován za supresor nádoru. V některých případech však může působit opačně onkogenně, protože působí jako promotor nádoru nebo stimulátor růstu,[16][69][70] pravděpodobně kvůli buněčným kontextům složeným z různých složek a aktivních drah. Typickým příkladem jsou pacienti s pankreatickým duktálním adenokarcinomem (PDAC), kteří mají mutace RBM10 a kteří vykazují míru přežití pozoruhodně vyšší než obecná míra přežití 5 let trvající PDAC méně než 7–8%.[50][71][72]
Paralogy a spojovací síť
RBM5 a RBM6 jsou paralogy RBM10. Byly generovány duplikací genů během evoluce genomu. Obecně fungují jako potlačující nádory[10][73][74][75][76][77][78][79] a jejich mutace jsou často identifikovány u rakoviny plic.[21] RBM5, RBM6 a RBM10 regulují alternativní sestřih[10][80][81] a obecně působí na různé RNA; v určitých případech však působí na stejnou podskupinu RNA a pravděpodobně mají synergické nebo antagonistické účinky.[10] Mezi RBM5 a RBM10 existuje křížová regulace; RBM10 snižuje úrovně transkriptu RBM5 pomocí alternativního sestřihu spojeného NMD.[14] Kromě toho rušení RBM10 (knockdown nebo nadměrná exprese) přináší změny sestřihu u více regulátorů sestřihu, včetně RBM5, a také významně ovlivňuje expresi jiných regulátorů sestřihu, včetně samotného RBM10.[9][14] Kromě toho jsou primární transkripty RBM10 podrobeny alternativnímu sestřihu na několika exonech neidentifikovanými regulátory sestřihu, což vede ke generování více izoforem RBM10. Tato data naznačují existenci alternativní sestřihové sítě tvořené RBM5, RBM6 a RBM10, jakož i dalšími regulátory sestřihu.[82] Očekává se, že studie na těchto sítích podpoří naše chápání transkriptomické homeostázy regulované spojováním a molekulárním a biologickým významem RBM10 v buňkách.
RBM10 reguluje stovky genů.[9][10][13][14][15] Další studie různých procesů a cest zprostředkovaných RBM10 mohou pomoci objasnit patogenezi a progresi onemocnění způsobených mutacemi RBM10 a mechanismy antitetických účinků RBM10 jako tumor supresoru a v některých případech jako promotoru nádoru a poskytnout vodítka pro lepší léčbu nemocí.
Reference
- ^ A b C GRCh38: Vydání souboru 89: ENSG00000182872 - Ensembl, Květen 2017
- ^ A b C GRCm38: Vydání souboru 89: ENSMUSG00000031060 - Ensembl, Květen 2017
- ^ „Human PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ „Myš PubMed Reference:“. Národní centrum pro biotechnologické informace, Americká národní lékařská knihovna.
- ^ Nagase T, Seki N, Tanaka A, Ishikawa K, Nomura N (srpen 1995). „Predikce kódujících sekvencí neidentifikovaných lidských genů. IV. Kódující sekvence 40 nových genů (KIAA0121-KIAA0160) odvozené analýzou cDNA klonů z lidské buněčné linie KG-1“. Výzkum DNA. 2 (4): 167–74, 199–210. doi:10.1093 / dnares / 2.4.167. PMID 8590280.
- ^ A b C Coleman MP, Ambrose HJ, Carrel L, Németh AH, Willard HF, Davies KE (leden 1996). „Nový gen, DXS8237E, leží do 20 kb proti proudu od UBE1 v Xp11.23 a má odlišný stav inaktivace X“. Genomika. 31 (1): 135–8. doi:10.1006 / geno.1996.0022. PMID 8808293.
- ^ A b C Thiselton DL, McDowall J, Brandau O, Ramser J, d'Esposito F, Bhattacharya SS a kol. (Duben 2002). „Integrovaná, funkčně anotovaná genová mapa intervalu DXS8026-ELK1 na lidském Xp11.3-Xp11.23: potenciální hotspot pro neurogenetické poruchy“. Genomika. 79 (4): 560–72. doi:10.1006 / geno.2002.6733. PMID 11944989.
- ^ Inoue A, Takahashi KP, Kimura M, Watanabe T, Morisawa S (srpen 1996). "Molekulární klonování proteinu vázajícího RNA, S1-1". Výzkum nukleových kyselin. 24 (15): 2990–7. doi:10.1093 / nar / 24.15.2990. PMC 146028. PMID 8760884.
- ^ A b C d E F G Wang Y, Gogol-Döring A, Hu H, Fröhler S, Ma Y, Jens M a kol. (Září 2013). „Integrativní analýza odhalila molekulární mechanismus, který je základem regulace spojování zprostředkovanou RBM10“. EMBO Molekulární medicína. 5 (9): 1431–42. doi:10,1002 / emmm.201302663. PMC 3799496. PMID 24000153.
- ^ A b C d E F G h i j k l m Bechara EG, Sebestyén E, Bernardis I, Eyras E, Valcárcel J (prosinec 2013). „RBM5, 6 a 10 odlišně regulují NUMB alternativní sestřih pro řízení proliferace rakovinných buněk“. Molekulární buňka. 52 (5): 720–33. doi:10.1016 / j.molcel.2013.11.010. PMID 24332178.
- ^ A b Inoue A, Yamamoto N, Kimura M, Nishio K, Yamane H, Nakajima K (březen 2014). "RBM10 reguluje alternativní sestřih". FEBS Dopisy. 588 (6): 942–7. doi:10.1016 / j.febslet.2014.01.052. PMID 24530524. S2CID 10303057.
- ^ Yang X, Coulombe-Huntington J, Kang S, Sheynkman GM, Hao T, Richardson A, et al. (Únor 2016). „Široké rozšíření schopností interakce proteinů alternativním sestřihem“. Buňka. 164 (4): 805–17. doi:10.1016 / j.cell.2016.01.029. PMC 4882190. PMID 26871637.
- ^ A b Sutherland LC, Thibault P, Durand M, Lapointe E, Knee JM, Beauvais A a kol. (Červenec 2017). „Sestřihová pole odhalují nové cíle RBM10, včetně pre-mRNA SMN2“. BMC Molekulární biologie. 18 (1): 19. doi:10.1186 / s12867-017-0096-x. PMC 5520337. PMID 28728573.
- ^ A b C d E Sun Y, Bao Y, Han W, Song F, Shen X, Zhao J a kol. (Srpen 2017). „Autoregulace RBM10 a křížová regulace RBM10 / RBM5 prostřednictvím alternativního sestřihu spojeného nesmyslu zprostředkovaného rozpadu“. Výzkum nukleových kyselin. 45 (14): 8524–8540. doi:10.1093 / nar / gkx508. PMC 5737846. PMID 28586478.
- ^ A b Collins KM, Kainov YA, Christodolou E, Ray D, Morris Q, Hughes T a kol. (Červen 2017). „Modul pro rozpoznávání RNA RRM-ZnF cílí na RBM10 na exonové sekvence, aby podpořil vyloučení exonu“. Výzkum nukleových kyselin. 45 (11): 6761–6774. doi:10.1093 / nar / gkx225. PMC 5499739. PMID 28379442.
- ^ A b C Loiselle JJ, Roy JG, Sutherland LC (2017). „RBM10 podporuje procesy spojené s transformací u malobuněčného karcinomu plic a je přímo regulován RBM5“. PLOS ONE. 12 (6): e0180258. Bibcode:2017PLoSO..1280258L. doi:10.1371 / journal.pone.0180258. PMC 5491171. PMID 28662214.
- ^ A b C d Johnston JJ, Teer JK, Cherukuri PF, Hansen NF, Loftus SK, Chong K a kol. (Květen 2010). „Masivně paralelní sekvenování exonů na chromozomu X identifikuje RBM10 jako gen, který způsobuje syndromickou formu rozštěpu patra“. American Journal of Human Genetics. 86 (5): 743–8. doi:10.1016 / j.ajhg.2010.04.007. PMC 2868995. PMID 20451169.
- ^ A b C Imielinski M, Berger AH, Hammerman PS, Hernandez B, Pugh TJ, Hodis E a kol. (Září 2012). "Mapování charakteristických znaků plicního adenokarcinomu s masivně paralelním sekvenováním". Buňka. 150 (6): 1107–20. doi:10.1016 / j.cell.2012.08.029. PMC 3557932. PMID 22980975.
- ^ A b C Seiler M, Peng S, Agrawal AA, Palacino J, Teng T, Zhu P a kol. (Duben 2018). „Somatická mutační krajina genů faktoru sestřihu a jejich funkční důsledky u 33 typů rakoviny“. Zprávy buněk. 23 (1): 282–296.e4. doi:10.1016 / j.celrep.2018.01.088. PMC 5933844. PMID 29617667.
- ^ A b Cieply B, Carstens RP (2015). „Funkční role alternativních sestřihových faktorů u lidských onemocnění“. Wiley Interdisciplinární hodnocení. RNA. 6 (3): 311–26. doi:10,1002 / wrna.1276. PMC 4671264. PMID 25630614.
- ^ A b Coomer AO, Black F, Greystoke A, Munkley J, Elliott DJ (2019). "Alternativní sestřih u rakoviny plic". Biochimica et Biophysica Acta (BBA) - regulační mechanismy genů. 1862 (11–12): 194388. doi:10.1016 / j.bbagrm.2019.05.006. PMID 31152916.
- ^ A b C Gorlin, RJ; Cervenka, J; Anderson, RC; Sauk, JJ; Bevis, WD (únor 1970). "Robinův syndrom. Pravděpodobně X-vázaná recesivní subvarieta vykazující perzistenci levé horní duté žíly a defekt síňového septa". American Journal of Diseases of Children (1960). 119 (2): 176–8. doi:10.1001 / archpedi.1970.02100050178020. PMID 5410571.
- ^ A b C d E Inoue A, Tsugawa K, Tokunaga K, Takahashi KP, Uni S, Kimura M a kol. (Září 2008). "Jaderné domény S1-1: charakterizace a dynamika jako funkce transkripční aktivity". Biologie buňky. 100 (9): 523–35. doi:10.1042 / BC20070142. PMID 18315527. S2CID 9893063.
- ^ A b Zheng S, Damoiseaux R, Chen L, Black DL (červen 2013). „Široce použitelná vysoce výkonná screeningová strategie identifikuje nové regulátory alternativního sestřihu Dlg4 (Psd-95)“. Výzkum genomu. 23 (6): 998–1007. doi:10,1101 / gr. 147546.112. PMC 3668367. PMID 23636947.
- ^ Lim J, Hao T, Shaw C, Patel AJ, Szabó G, Rual JF a kol. (Květen 2006). „Síť interakce protein-protein pro lidské zděděné ataxie a poruchy degenerace Purkyňových buněk“. Buňka. 125 (4): 801–14. doi:10.1016 / j.cell.2006.03.032. PMID 16713569. S2CID 13709685.
- ^ Mohan N, Kumar V, Kandala DT, Kartha CC, Laishram RS (září 2018). "Funkce nezávislá na sestřihu RBM10 řídí specifické 3 'zpracování UTR k regulaci srdeční hypertrofie". Zprávy buněk. 24 (13): 3539–3553. doi:10.1016 / j.celrep.2018.08.077. PMID 30257214.
- ^ A b C Mueller CF, Berger A, Zimmer S, Tiyerili V, Nickenig G (srpen 2009). „Heterogenní jaderný riboprotein S1-1 reguluje expresi genu receptoru AT1 prostřednictvím transkripčních a posttranskripčních mechanismů“. Archivy biochemie a biofyziky. 488 (1): 76–82. doi:10.1016 / j.abb.2009.06.002. PMID 19508861.
- ^ Treiber T, Treiber N, Plessmann U, Harlander S, Daiß JL, Eichner N a kol. (Duben 2017). „Kompendium proteinů vázajících RNA, které regulují biogenezi mikroRNA“. Molekulární buňka. 66 (2): 270–284.e13. doi:10.1016 / j.molcel.2017.03.014. PMID 28431233.
- ^ A b C Jung JH, Lee H, Cao B, Liao P, Zeng SX, Lu H (leden 2020). "RNA-vazebný motivový protein 10 indukuje apoptózu a potlačuje proliferaci aktivací p53". Onkogen. 39 (5): 1031–1040. doi:10.1038 / s41388-019-1034-9. PMC 6994357. PMID 31591476.
- ^ Guan G, Li R, Tang W, Liu T, Su Z, Wang Y a kol. (Březen 2017). „Exprese motivu vázajícího RNA 10 je spojena s pokročilým stadiem nádoru a maligním chováním buněk rakoviny plicního adenokarcinomu.“. Biologie nádorů. 39 (3): 1010428317691740. doi:10.1177/1010428317691740. PMID 28347232. S2CID 206612545.
- ^ Kunimoto H, Inoue A, Kojima H, Yang J, Zhao H, Tsuruta D, Nakajima K (únor 2020). „RBM10 reguluje duplikaci centriolů v buňkách HepG2 ektopickým shromažďováním komplexů PLK4-STIL v jádře“. Geny do buněk. 25 (2): 100–110. doi:10.1111 / gtc.12741. PMID 31820547. S2CID 209165475.
- ^ Pozzi B, Bragado L, Mammi P, Torti MF, Gaioli N, Gebhard LG a kol. (Červenec 2020). „Virus dengue cílí na RBM10 deregulující sestřih hostitelských buněk a vrozenou imunitní odpověď“. Výzkum nukleových kyselin. 48 (12): 6824–6838. doi:10.1093 / nar / gkaa340. PMC 7337517. PMID 32432721.
- ^ Salichs E, Ledda A, Mularoni L, Albà MM, de la Luna S (březen 2009). „Analýza genitů opakujících se histidinu v celém genomu odhaluje jejich roli v lokalizaci lidských proteinů do kompartmentu jaderných skvrn“. Genetika PLOS. 5 (3): e1000397. doi:10.1371 / journal.pgen.1000397. PMC 2644819. PMID 19266028.
- ^ Goto Y, Kimura H (prosinec 2009). „Neaktivní modifikace histonů H3 specifických pro chromozomy X a hypometylace CpG ohraničují hranici chromatinu mezi X-inaktivovaným a únikovým genem“. Výzkum nukleových kyselin. 37 (22): 7416–28. doi:10.1093 / nar / gkp860. PMC 2794193. PMID 19843608.
- ^ Stes E, Laga M, Walton A, Samyn N, Timmerman E, De Smet I a kol. (Červen 2014). "Protokol COFRADIC ke studiu ubikvitinace proteinů". Journal of Proteome Research. 13 (6): 3107–13. doi:10.1021 / pr4012443. PMID 24816145.
- ^ Akimov V, Barrio-Hernandez I, Hansen SV, Hallenborg P, Pedersen AK, Bekker-Jensen DB a kol. (Červenec 2018). "UbiSite přístup pro komplexní mapování lysinu a N-terminálních ubikvitinačních míst". Přírodní strukturní a molekulární biologie. 25 (7): 631–640. doi:10.1038 / s41594-018-0084-r. PMID 29967540. S2CID 49559977.
- ^ Choudhary C, Kumar C, Gnad F, Nielsen ML, Rehman M, Walther TC a kol. (Srpen 2009). „Lysinová acetylace je zaměřena na proteinové komplexy a reguluje hlavní buněčné funkce“. Věda. 325 (5942): 834–40. Bibcode:2009Sci ... 325..834C. doi:10.1126 / science.1175371. PMID 19608861. S2CID 206520776.
- ^ Guo A, Gu H, Zhou J, Mulhern D, Wang Y, Lee KA a kol. (Leden 2014). "Obohacení imunoafinitou a hmotnostní spektrometrická analýza methylace proteinů". Molekulární a buněčná proteomika. 13 (1): 372–87. doi:10,1074 / mcp.O113.027870. PMC 3879628. PMID 24129315.
- ^ Network Atlas Research Network pro rakovinu (červenec 2014). "Komplexní molekulární profil adenokarcinomu plic". Příroda. 511 (7511): 543–50. Bibcode:2014Natur.511..543T. doi:10.1038 / příroda13385. PMC 4231481. PMID 25079552.
- ^ Yuan Y, Liu L, Chen H, Wang Y, Xu Y, Mao H a kol. (Květen 2016). „Komplexní charakterizace molekulárních rozdílů v rakovině mezi pacienty a pacientkami“. Rakovinová buňka. 29 (5): 711–722. doi:10.1016 / j.ccell.2016.04.001. PMC 4864951. PMID 27165743.
- ^ Yin LL, Wen XM, Li M, Xu YM, Zhao XF, Li J, Wang XW (listopad 2018). „Genová mutace v proteinu 10 vázajícím RNA je spojena s progresí plicního adenokarcinomu a špatnou prognózou“. Onkologické dopisy. 16 (5): 6283–6292. doi:10.3892 / ol.2018.9496. PMC 6202477. PMID 30405763.
- ^ Powis, Z; Hart, A; Cherny, S; Petrik, já; Palmaer, E; Tang, S; Jones, C (2. června 2017). „Klinické diagnostické hodnocení exomu u kojence se smrtelnou poruchou: genetická diagnostika syndromu TARP a rozšíření fenotypu u pacienta s nově hlášenou změnou RBM10“. Lékařská genetika BMC. 18 (1): 60. doi:10.1186 / s12881-017-0426-3. PMC 5455125. PMID 28577551.
- ^ Gripp KW, Hopkins E, Johnston JJ, Krause C, Dobyns WB, Biesecker LG (říjen 2011). „Dlouhodobé přežití u syndromu TARP a potvrzení RBM10 jako genu způsobujícího onemocnění“. American Journal of Medical Genetics. Část A. 155A (10): 2516–20. doi:10,1002 / ajmg.a.34190. PMC 3183328. PMID 21910224.
- ^ Niceta M, Barresi S, Pantaleoni F, Capolino R, Dentici ML, Ciolfi A a kol. (Červen 2019). „TARP syndrom: Dlouhodobé přežití, anatomické vzorce vrozených srdečních vad, diferenciální diagnostika a patogenetické úvahy“. European Journal of Medical Genetics. 62 (6): 103534. doi:10.1016 / j.ejmg.2018.09.001. PMID 30189253.
- ^ Højland AT, Lolas I, Okkels H, Lautrup CK, Diness BR, Petersen MB, Nielsen IK (prosinec 2018). „První hlášený dospělý pacient se syndromem TARP: kazuistika“. American Journal of Medical Genetics. Část A. 176 (12): 2915–2918. doi:10,1002 / ajmg.a. 40638. PMC 6587983. PMID 30462380.
- ^ Loiselle JJ, Sutherland LC (květen 2018). „RBM10: škodlivé nebo užitečné - je třeba vzít v úvahu mnoho faktorů. Journal of Cellular Biochemistry. 119 (5): 3809–3818. doi:10.1002 / jcb.26644. PMC 5901003. PMID 29274279.
- ^ Xia QY, Wang XT, Zhan XM, Tan X, Chen H, Liu Y a kol. (Květen 2017). „Translokační karcinomy ledvinových buněk (RCC) s Xp11 s genovou fúzí RBM10-TFE3 prokazující melanotické rysy a překrývající se morfologii s t (6; 11) RCC: Zájem a diagnostická úskalí při detekci paracentrické inverze TFE3“. American Journal of Surgical Pathology. 41 (5): 663–676. doi:10.1097 / PAS.0000000000000837. PMID 28288037. S2CID 205918230.
- ^ Argani P, Zhang L, Reuter VE, Tickoo SK, Antonescu CR (květen 2017). „RBM10-TFE3 karcinom ledvinových buněk: potenciální diagnostická úskalí v důsledku kryptické intrachromozomální inverze Xp11.2 vedoucí k falešně negativní TFE3 FISH“. American Journal of Surgical Pathology. 41 (5): 655–662. doi:10.1097 / PAS.0000000000000835. PMC 5391276. PMID 28296677.
- ^ Kato I, Furuya M, Baba M, Kameda Y, Yasuda M, Nishimoto K a kol. (Srpen 2019). „RBM10-TFE3 karcinom z renálních buněk charakterizovaný paracentrickou inverzí s konzistentně úzce rozdělenými signály při rozpadové fluorescenční hybridizaci in situ: studie 10 případů a přehled literatury“. Histopatologie. 75 (2): 254–265. doi:10.1111 / his.13866. PMID 30908700. S2CID 85516169.
- ^ A b Witkiewicz AK, McMillan EA, Balaji U, Baek G, Lin WC, Mansour J a kol. (Duben 2015). „Celoexomové sekvenování rakoviny pankreatu definuje genetickou rozmanitost a terapeutické cíle“. Příroda komunikace. 6: 6744. Bibcode:2015NatCo ... 6,6744 W.. doi:10.1038 / ncomms7744. PMC 4403382. PMID 25855536.
- ^ Furukawa T, Kuboki Y, Tanji E, Yoshida S, Hatori T, Yamamoto M a kol. (2011). „Sekvenování celého exomu odhaluje časté mutace GNAS v intraduktálních papilárních mucinózních novotvarech slinivky břišní“. Vědecké zprávy. 1: 161. Bibcode:2011NatSR ... 1E.161F. doi:10.1038 / srep00161. PMC 3240977. PMID 22355676.
- ^ Giannakis M, Mu XJ, Shukla SA, Qian ZR, Cohen O, Nishihara R a kol. (Duben 2016). „Genomické koreláty infiltrátů imunitních buněk v kolorektálním karcinomu“. Zprávy buněk. 15 (4): 857–865. doi:10.1016 / j.celrep.2016.03.075. PMC 4850357. PMID 27149842.
- ^ Lawrence MS, Stojanov P, Mermel CH, Robinson JT, Garraway LA, Golub TR a kol. (Leden 2014). „Analýza objevů a saturace rakovinových genů u 21 typů nádorů“. Příroda. 505 (7484): 495–501. Bibcode:2014 Natur.505..495L. doi:10.1038 / příroda12912. PMC 4048962. PMID 24390350.
- ^ Ibrahimpasic T, Xu B, Landa I, Dogan S, Middha S, Seshan V a kol. (Říjen 2017). „RBM10 jako nové geny rakoviny štítné žlázy spojené s virulencí nádoru“. Klinický výzkum rakoviny. 23 (19): 5970–5980. doi:10.1158 / 1078-0432.CCR-17-1183. PMC 5626586. PMID 28634282.
- ^ Antonello ZA, Hsu N, Bhasin M, Roti G, Joshi M, Van Hummelen P a kol. (Říjen 2017). „V600E“. Cílový cíl. 8 (49): 84743–84760. doi:10,18632 / oncotarget.21262. PMC 5689570. PMID 29156680.
- ^ Ibrahimpasic T, Ghossein R, Shah JP, Ganly I (březen 2019). „Špatně diferencovaný karcinom štítné žlázy: současný stav a budoucí vyhlídky“. Štítná žláza. 29 (3): 311–321. doi:10.1089 / tvůj.2018.0509. PMC 6437626. PMID 30747050.
- ^ A b Kan Z, Jaiswal BS, Stinson J, Janakiraman V, Bhatt D, Stern HM a kol. (Srpen 2010). „Rozmanité vzorce somatických mutací a změny dráhy u lidských rakovin“. Příroda. 466 (7308): 869–73. Bibcode:2010Natur.466..869K. doi:10.1038 / nature09208. PMC 3026267. PMID 20668451.
- ^ Tian W, Hu W, Shi X, Liu P, Ma X, Zhao W a kol. (Duben 2020). „Komplexní genomický profil cholangiokarcinomů v Číně“. Onkologické dopisy. 19 (4): 3101–3110. doi:10,3892 / ol.2020.11429. PMC 7074170. PMID 32256810.
- ^ Schwab ME, Song H, Mattis A, Phelps A, Vu LT, Huang FW, Nijagal A (březen 2020). „Somatické mutace de novo a amplifikace KRAS jsou spojeny s cholangiokarcinomem u pacienta s anamnézou choledochální cysty“. Journal of Pediatric Surgery. doi:10.1016 / j.jpedsurg.2020.03.008. PMID 32295706.
- ^ Juratli TA, McCabe D, Nayyar N, Williams EA, Silverman IM, Tummala SS a kol. (Listopad 2018). „DMD genomové delece charakterizují podmnožinu progresivních / vyšších meningiomů se špatným výsledkem“. Acta Neuropathologica. 136 (5): 779–792. doi:10.1007 / s00401-018-1899-7. PMID 30123936. S2CID 52039057.
- ^ Majd NK, Metrus NR, Santos-Pinheiro F, Trevino CR, Fuller GN, Huse JT a kol. (Únor 2019). "Zkrácení RBM10 v astroblastomu u pacienta s anamnézou mandibulárního ameloblastomu: kazuistika". Genetika rakoviny. 231-232: 41–45. doi:10.1016 / j.cancergen.2019.01.001. PMID 30803556.
- ^ Misquitta-Ali CM, Cheng E, O'Hanlon D, Liu N, McGlade CJ, Tsao MS, Blencowe BJ (leden 2011). „Globální profilování a molekulární charakterizace alternativních sestřihových událostí nesprávně regulovaných u rakoviny plic“. Molekulární a buněčná biologie. 31 (1): 138–50. doi:10.1128 / MCB.00709-10. PMC 3019846. PMID 21041478.
- ^ A b Hernández J, Bechara E, Schlesinger D, Delgado J, Serrano L, Valcárcel J (2016). "Vlastnosti potlačující nádory regulačního faktoru sestřihu RBM10". RNA Biology. 13 (4): 466–72. doi:10.1080/15476286.2016.1144004. PMC 4841610. PMID 26853560.
- ^ A b C Zhao J, Sun Y, Huang Y, Song F, Huang Z, Bao Y a kol. (Leden 2017). „Funkční analýza odhaluje, že mutace RBM10 přispívají k patogenezi adenokarcinomu plic deregulací sestřihu“. Vědecké zprávy. 7: 40488. Bibcode:2017NatSR ... 740488Z. doi:10.1038 / srep40488. PMC 5238425. PMID 28091594.
- ^ A b Han LP, Wang CP, Han SL (říjen 2018). „Nadměrná exprese RBM10 indukuje apoptózu buněk osteosarkomu a inhibuje buněčnou proliferaci a migraci“. Médecine / Sciences. 34 Zaostřeno na F1: 81–86. doi:10.1051 / medsci / 201834f114. PMID 30403180.
- ^ Jin X, Di X, Wang R, Ma H, Tian C, Zhao M a kol. (Červen 2019). „RBM10 inhibuje buněčnou proliferaci adenokarcinomu plic prostřednictvím signální dráhy RAP1 / AKT / CREB“. Journal of Cellular and Molecular Medicine. 23 (6): 3897–3904. doi:10,1111 / jcmm.14263. PMC 6533519. PMID 30955253.
- ^ Sutherland LC, Rintala-Maki ND, White RD, Morin CD (leden 2005). „Proteiny vázající RNA (RBM): nová rodina modulátorů apoptózy?“. Journal of Cellular Biochemistry. 94 (1): 5–24. doi:10.1002 / jcb.20204. PMID 15514923. S2CID 26344717.
- ^ Wang K, Bacon ML, Tessier JJ, Rintala-Maki ND, Tang V, Sutherland LC (2012). „RBM10 moduluje apoptózu a ovlivňuje expresi genu TNF-α“. Journal of Cell Death. 5: 1–19. doi:10,4137 / JCD.S9073. PMC 4583097. PMID 26446321.
- ^ Rodor J, FitzPatrick DR, Eyras E, Cáceres JF (leden 2017). „Prostředí vázající RNA RBM10 a jeho role v regulaci alternativního sestřihu v modelech raného vývoje myší“. RNA Biology. 14 (1): 45–57. doi:10.1080/15476286.2016.1247148. PMC 5270529. PMID 27763814.
- ^ Sun X, Jia M, Sun W, Feng L, Gu C, Wu T (únor 2019). "Funkční role RBM10 v proliferaci adenokarcinomu plic". International Journal of Oncology. 54 (2): 467–478. doi:10.3892 / ijo.2018.4643. PMC 6317669. PMID 30483773.
- ^ Balachandran VP, Łuksza M, Zhao JN, Makarov V, Moral JA, Remark R a kol. (Listopad 2017). „Identifikace jedinečných neoantigenových kvalit u dlouhodobě přeživších pacientů s rakovinou pankreatu“. Příroda. 551 (7681): 512–516. Bibcode:2017Natur.551..512B. doi:10.1038 / příroda24462. PMC 6145146. PMID 29132146.
- ^ Siegel RL, Miller KD, Jemal A (leden 2018). „Statistics oncer, 2018“. Ca.. 68 (1): 7–30. doi:10,3322 / caac.21442. PMID 29313949.
- ^ Mourtada-Maarabouni M, Williams GT (červenec 2002). „RBM5 / LUCA-15 - potlačení nádoru kontrolou apoptózy a buněčného cyklu?“. TheScientificWorldJournal. 2: 1885–90. doi:10.1100 / tsw.2002.859. PMC 6009235. PMID 12920317.
- ^ Oh JJ, Razfar A, Delgado I, Reed RA, Malkina A, Boctor B, Slamon DJ (duben 2006). „3p21.3 tumor supresorový gen H37 / Luca15 / RBM5 inhibuje růst lidských buněk rakoviny plic zastavením buněčného cyklu a apoptózou“. Výzkum rakoviny. 66 (7): 3419–27. doi:10.1158 / 0008-5472.CAN-05-1667. PMID 16585163.
- ^ Fushimi K, Ray P, Kar A, Wang L, Sutherland LC, Wu JY (říjen 2008). "Up-regulace proapoptotické sestřihové izoformy kaspázy 2 kandidátem potlačujícím nádor, RBM5". Sborník Národní akademie věd Spojených států amerických. 105 (41): 15708–13. Bibcode:2008PNAS..10515708F. doi:10.1073 / pnas.0805569105. PMC 2572934. PMID 18840686.
- ^ Bonnal S, Martínez C, Förch P, Bachi A, Wilm M, Valcárcel J (říjen 2008). "RBM5 / Luca-15 / H37 reguluje párování alternativních sestřihových míst Fas po definici exonu". Molekulární buňka. 32 (1): 81–95. doi:10.1016 / j.molcel.2008.08.008. PMID 18851835.
- ^ Sutherland LC, Wang K, Robinson AG (březen 2010). "RBM5 jako předpokládaný gen potlačující nádor pro rakovinu plic". Journal of Thoracic Oncology. 5 (3): 294–8. doi:10.1097 / JTO.0b013e3181c6e330. PMID 20186023.
- ^ Jamsai D, Watkins DN, O'Connor AE, Merriner DJ, Gursoy S, Bird AD a kol. (Listopad 2017). „Důkazy in vivo, že RBM5 je nádorový supresor v plicích“. Vědecké zprávy. 7 (1): 16323. Bibcode:2017NatSR ... 716323J. doi:10.1038 / s41598-017-15874-9. PMC 5701194. PMID 29176597.
- ^ Wang Q, Wang F, Zhong W, Ling H, Wang J, Cui J a kol. (Květen 2019). „Protein vázající RNA RBM6 jako tumor supresorový gen potlačuje růst a progresi v laryngokarcinomu“. Gen. 697: 26–34. doi:10.1016 / j.gene.2019.02.025. PMID 30772516.
- ^ Deckert J, Hartmuth K, Boehringer D, Behzadnia N, Will CL, Kastner B a kol. (Červenec 2006). "Proteinové složení a struktura elektronové mikroskopie afinitně purifikovaných lidských spliceosomálních B komplexů izolovaných za fyziologických podmínek". Molekulární a buněčná biologie. 26 (14): 5528–43. doi:10.1128 / MCB.00582-06. PMC 1592722. PMID 16809785.
- ^ Papasaikas P, Tejedor JR, Vigevani L, Valcárcel J (leden 2015). „Funkční spojovací síť odhaluje rozsáhlý regulační potenciál jádra spliceozomálního aparátu“. Molekulární buňka. 57 (1): 7–22. doi:10.1016 / j.molcel.2014.10.030. PMID 25482510. S2CID 11133534.
- ^ Ule J, Blencowe BJ (říjen 2019). "Alternativní sestřih regulačních sítí: funkce, mechanismy a vývoj". Molekulární buňka. 76 (2): 329–345. doi:10.1016 / j.molcel.2019.09.017. PMID 31626751.
Další čtení
- Maruyama K, Sugano S (leden 1994). „Oligo-capping: jednoduchá metoda k nahrazení struktury cap eukaryotických mRNA oligoribonukleotidy“. Gen. 138 (1–2): 171–4. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (říjen 1997). "Konstrukce a charakterizace knihovny cDNA obohacené o celou délku a 5'-end". Gen. 200 (1–2): 149–56. doi:10.1016 / S0378-1119 (97) 00411-3. PMID 9373149.
- Hartley JL, Temple GF, Brasch MA (listopad 2000). „Klonování DNA pomocí in vitro místně specifické rekombinace“. Výzkum genomu. 10 (11): 1788–95. doi:10,1101 / gr. 143000. PMC 310948. PMID 11076863.
- Wiemann S, Weil B, Wellenreuther R, Gassenhuber J, Glassl S, Ansorge W a kol. (Březen 2001). „Směrem ke katalogu lidských genů a proteinů: sekvenování a analýza 500 nových kompletních proteinů kódujících lidské cDNA“. Výzkum genomu. 11 (3): 422–35. doi:10,1101 / gr. GR1547R. PMC 311072. PMID 11230166.
- Li J, Hawkins IC, Harvey CD, Jennings JL, Link AJ, Patton JG (listopad 2003). „Regulace alternativního sestřihu pomocí SRrp86 a jeho interagujících proteinů“. Molekulární a buněčná biologie. 23 (21): 7437–47. doi:10.1128 / MCB.23.21.7437-7447.2003. PMC 207616. PMID 14559993.
- Beausoleil SA, Jedrychowski M, Schwartz D, Elias JE, Villén J, Li J a kol. (Srpen 2004). „Rozsáhlá charakterizace jaderných fosfoproteinů z buněk HeLa“. Sborník Národní akademie věd Spojených států amerických. 101 (33): 12130–5. Bibcode:2004PNAS..10112130B. doi:10.1073 / pnas.0404720101. PMC 514446. PMID 15302935.
- Ballif BA, Villén J, Beausoleil SA, Schwartz D, Gygi SP (listopad 2004). „Fosfoproteomická analýza vyvíjejícího se myšího mozku“. Molekulární a buněčná proteomika. 3 (11): 1093–101. doi:10,1074 / mcp.M400085-MCP200. PMID 15345747.
- Wiemann S, Arlt D, Huber W, Wellenreuther R, Schleeger S, Mehrle A a kol. (Říjen 2004). „Od ORFeome k biologii: funkční plynovod genomiky“. Výzkum genomu. 14 (10B): 2136–44. doi:10,1101 / gr. 2576704. PMC 528930. PMID 15489336.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N a kol. (Říjen 2005). „Směrem k mapě lidské interakční sítě protein-protein v měřítku proteomu“. Příroda. 437 (7062): 1173–8. Bibcode:2005 Natur.437.1173R. doi:10.1038 / nature04209. PMID 16189514. S2CID 4427026.
- Mehrle A, Rosenfelder H, Schupp I, del Val C, Arlt D, Hahne F a kol. (Leden 2006). „Databáze LIFEdb v roce 2006“. Výzkum nukleových kyselin. 34 (Problém s databází): D415-8. doi:10.1093 / nar / gkj139. PMC 1347501. PMID 16381901.
- Martínez-Arribas F, Agudo D, Pollán M, Gómez-Esquer F, Díaz-Gil G, Lucas R, Schneider J (duben 2006). „Pozitivní korelace mezi expresí genů RBM chromozomu X (RBMX, RBM3, RBM10) a proapoptotickým genem Bax u lidské rakoviny prsu“. Journal of Cellular Biochemistry. 97 (6): 1275–82. doi:10.1002 / jcb.20725. PMID 16552754. S2CID 9804734.
- Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (listopad 2006). „Globální, in vivo a místně specifická dynamika fosforylace v signálních sítích“. Buňka. 127 (3): 635–48. doi:10.1016 / j.cell.2006.09.026. PMID 17081983. S2CID 7827573.