Polysome profilování - Polysome profiling

Polysome profilování je technika v molekulární biologie který se používá ke studiu asociace mRNA s ribozomy. Je důležité si uvědomit, že tato technika se liší od profilování ribozomu. Obě techniky byly přezkoumány[1] a oba se používají při analýze translatome, ale data, která generují, jsou na velmi odlišné úrovni specificity. Při použití odborníky je technika pozoruhodně reprodukovatelná: 3 profily v prvním snímku pocházejí ze 3 různých experimentů.[2]
Postup
Postup začíná vytvořením a buněčný lyzát sledovaných buněk. Tento lyzát obsahuje polysomy monosomy (složené z jednoho ribozomu pobývajícího na mRNA ), malý (40S v eukaryoty ) a velké (60S u eukaryot) ribozomální podjednotky, „volná“ mRNA a řada dalších rozpustný buněčné komponenty.
Postup pokračuje vytvořením kontinuity sacharóza gradient spojitě proměnné hustoty v a odstředivka trubka. Při použitých koncentracích (15-45% v příkladu) sacharóza nenarušuje asociaci ribozomů a mRNA. 15% část gradientu je v horní části trubice, zatímco 45% část je dole kvůli jejich odlišnosti hustota.
Konkrétní částka (měřeno pomocí optická hustota ) lyzátu se pak jemně navrství na gradient v trubici. Lyzát, i když obsahuje velké množství rozpustného materiálu, je mnohem méně hustý než 15% sacharózy, a proto jej lze udržovat jako samostatnou vrstvu v horní části zkumavky, pokud je to provedeno šetrně.
Za účelem oddělení složek lyzátu se přípravek podrobí odstředění. Tento zrychluje složky lyzátu mnohonásobně gravitační síla a tak je pohání přes gradient na základě toho, jak „velké“ jsou jednotlivé komponenty. Malé (40S) podjednotky cestují do přechodu méně daleko než velké (60S) podjednotky. Ribsomy 80S na mRNA se pohybují dále (všimněte si, že příspěvek velikosti mRNA k ujeté vzdálenosti není významný). Polysomy složené ze 2 ribozomů se pohybují dále, polysomy se 3 ribozomy se pohybují ještě dále a dál a dál. "Velikost" komponent je označena S, svedberg jednotka. Všimněte si, že jeden S = 10−13 sekund a že pojem „velký“ je ve skutečnosti zjednodušením.

Po centrifugaci se obsah zkumavky sbírá jako frakce shora (menší, pomalejší) dolů (větší, rychlejší) a stanoví se optická hustota frakcí. První odstraněné frakce mají velké množství relativně malých molekul, jako jsou tRNA, jednotlivé proteiny atd.
Aplikace
Tuto techniku je možné použít ke studiu celkového stupně translace v buňkách (například[3][4][5]), ale lze jej mnohem konkrétněji použít ke studiu jednotlivých proteinů a jejich mRNA. Jako příklad zobrazený ve spodní části obrázku může být protein, který tvoří část malé podjednotky, nejprve detekován ve frakci 40S, poté téměř zmizí z frakce 60S (separace na těchto gradientech nejsou absolutní), poté se znovu objeví ve frakcích 80S a polysomech. To naznačuje, že v buňce se nachází maximálně velmi málo proteinu, který není součástí malé podjednotky. Naproti tomu v horní řadě obrázku imunoblotu se rozpustný protein objevuje v rozpustných frakcích a je asociován s ribozomy a polysomy. Konkrétní protein je a chaperonový protein, který (ve zkratce) pomáhá skládat rodící se peptid protože je vytlačován z ribozomu. Jako jiná práce
v článku, který ukázal, existuje přímá asociace chaperonu s ribozomem.[2]
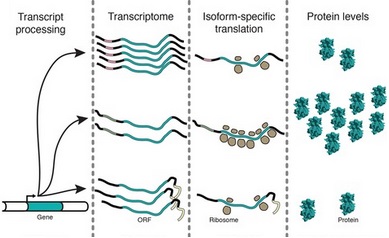
Tato technika může být také použita ke studiu stupně translace konkrétní mRNA[6] V těchto experimentech byly 5 'a 3' sekvence mRNA zkoumány z hlediska jejich účinků na množství produkované mRNA a jak dobře byly mRNA přeloženy. Jak je znázorněno, ne všechny izoformy mRNA jsou přeloženy se stejnou účinností Přestože jejich kódovací sekvence jsou stejné.[6]
Reference
- ^ Piccirillo, CA; et al. (2014). "Translační kontrola imunitních odpovědí: od transkriptů po translatomy". Přírodní imunologie. 15 (6): 503–511. doi:10.1038 / ni.2891. PMID 24840981.
- ^ A b Hanebuth, MA; et al. (2016). „Multivalentní kontakty Hsp70 Ssb přispívají k jeho architektuře na ribozomech a rodící se řetězové interakci“. Příroda komunikace. 7: 13695. Bibcode:2016NatCo ... 713695H. doi:10.1038 / ncomms13695. PMC 5150220. PMID 27917864.
- ^ Lin, CJ; et al. (2010). „Antidepresivum sertralin inhibuje iniciaci translace omezením savčího cíle signalizace rapamycinem“. Výzkum rakoviny. 70 (8): 3199–3208. doi:10.1158 / 0008-5472.CAN-09-4072. PMID 20354178.
- ^ Coudert, L; et al. (2014). "Analýza iniciace translace během stresových podmínek pomocí polysomového profilování". Žurnál vizualizovaných experimentů (87). doi:10.3791/51164. PMC 4193336. PMID 24893838.
- ^ Molon, M; et al. (2016). „Rychlost metabolismu jako faktor určující dlouhověkost kvasinek Saccharomyces cerevisiae“. Věk (Dordrecht, Nizozemsko). 38 (1): 11. doi:10.1007 / s11357-015-9868-8. PMC 5005888. PMID 26783001.
- ^ A b Patro, SN; Doudna, JA (2016). "Laditelná syntéza bílkovin transkriptovými izoformami v lidských buňkách". eLife. 5. doi:10.7554 / eLife.10921. PMC 4764583. PMID 26735365.