Mad1 - Mad1 - Wikipedia
| Mad1 | |||||||
|---|---|---|---|---|---|---|---|
 Krystalová struktura, tetramer komplexu Mad1-Mad2, žlutá a červená = monomery Mad1, bledozelená = monomery Mad2 | |||||||
| Identifikátory | |||||||
| Organismus | |||||||
| Symbol | MAD1 | ||||||
| Entrez | 852794 | ||||||
| PDB | 1GO4 | ||||||
| RefSeq (mRNA) | NM_001180951.3 | ||||||
| RefSeq (Prot) | NP_011429.3 | ||||||
| UniProt | P40957 | ||||||
| Další údaje | |||||||
| Chromozóm | VII: 0,35 - 0,35 Mb | ||||||
| |||||||
Mad1 je nepodstatné protein v kterém droždí má funkci v kontrolní bod sestavy vřetena (VAK).[1]Tento kontrolní bod monitoruje připojení chromozomů k mikrotubulům vřetena a brání buňkám ve spouštění anafáze, dokud se vřeteno nevytvoří. Název Mad odkazuje na pozorování, že mutantní buňky mají během depolymerace mikrotubulů deficit mitotické zástavy (MAD). Mad1 rekrutuje inhibitor anafáze Mad2 k nepřipojeným kinetochorům a je nezbytný pro Mad2-CDC20 komplexní formace in vivo ale ne in vitro. In vivo„Mad1 působí jako kompetitivní inhibitor komplexu Mad2-Cdc20.[2] Mad1 je fosforylován Mps1, který pak vede společně s dalšími aktivitami k tvorbě komplexu mitotického kontrolního bodu (MCC). Tím inhibuje aktivitu komplexu / cyklosomu podporujícího anafázu (APC / C). Homology Mad1 jsou konzervovány v eukaryotech od kvasinek po savce.
Úvod
Na počátku 90. let byly identifikovány geny kvasinek, jejichž mutace vedla k defektu mitotické zástavy v reakci na rozebrání mikrotubulů (geny s deficitem mitotické zástavy - geny MAD). Tyto buňky nevykazovaly žádnou mitotickou zástavu v přítomnosti inhibitorů polymerace mikrotubulů, a proto nebyly schopny oddálit dělení buněk.[1] Identifikované geny zahrnovaly MAD1, MAD2 a MAD3 geny. Ve všech jsou zachovány eukaryoty a jsou zapojeni do cesty, která je aktivní v prometafáze zabránit předčasnému odloučení sestry chromatidy a tvoří tzv kontrolní bod sestavy vřetena (VAK). Tento kontrolní bod sleduje stav připojení chromozomů k mitotickému vřetenu a inhibuje metafáze na anafáze přechod tím, že zabrání aktivaci komplex podporující anafázi / cyklosom (APC / C), a tím i degradace z buněčný cyklus regulátory.[3] Mad1 je v této dráze akumulován v nepřipojených kinetochorech a funguje jako senzor pro nepřipojené kinetochory v tomto strojním zařízení.
Funkce
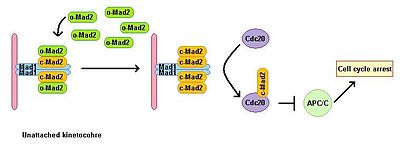
Eukaryotické buňky vykazují mitotickou zástavu v přítomnosti inhibitorů polymerace mikrotubulů. Kontrolní bod sestavy vřetena sleduje stav vřetena a spojuje přechod metafáze-anafáze se správným bipolárním připojením všech kinetochorů k mitotickému vřetenu. Kontrolní bod sestavy vřetena inhibuje aktivitu komplexu podporujícího anafázu tím, že brání degradaci následných efektorů, což by jinak vedlo k nástupu a výstupu anafáze z mitózy. Vyčerpání Mad1 vede ke ztrátě VAK funkce. Mad1 se lokalizuje převážně na nepřipojených kinetochorech a spouští mitotickou zástavu v případě jediného nepřipojeného kinetochore. Mad1 získává důležitou součást SAC Mad2 na nepřipojené kinetochory a indukuje zesílení signálu mitotické zástavy. Existuje skupina volného cytoplazmatického Mad2 v jeho neaktivní otevřené konformaci zvané o-MAD2. Když je vázán na Mad1, Mad2 přijme aktivní konformace nazývá uzavřené (c-Mad2) a tvoří heterotetramer dvou jednotek Mad1 a dvou jednotek c-Mad2. Heterotetramer Mad1 – c-Mad2 je velmi stabilní a funguje jako katalytické receptor zdarma cytoplazmatický o-Mad2. Volný o-Mad2 se váže na tento receptor a mění jeho konformaci na aktivní uzavřenou formu. Tento druhý c-MAD2 je přenesen do CDC20 se dosud neznámým mechanismem a tvoří komplex Cdc20 – c-Mad2. Tento komplex je základní součástí komplexu mitotického kontrolního bodu (MCC). MCC se váže a inhibuje APC / C a proto zastaví progresi mitózou.[3][4]
Nařízení
Jsou dva kontrolní body proti proudu kinázy podílí se na regulaci funkce Mad1 prostřednictvím fosforylace.[5] Mps1 fosforyluje Mad1 oba in vitro a in vivo a předpokládá se, že reguluje lokalizaci Mad1 a Mad2 na kinetochory a jejich dynamika interakce. BUB1 je další kináza, která rekrutuje Mad1 do kinetochorů a aktivuje ji, pokud je kinetochor nepřipojen.[3]Pokud je k vřetenu připojen kinetochor, inhibitor SAC p31kometa inhibuje Mad1 zprostředkované konformační přeskupení Mad2 a zabraňuje Mad2 ve vazbě na Cdc20.[3]
Strukturální prvky a mechanismus

Podle biochemických metod se předpokládalo, že Mad1 kóduje 90kD, zbytek 718,[6] svinutá cívka protein s charakteristickým tvarem tyče[1]v roce 1995. Krystalové struktury brzy následovaly. V roce 2002 byla zveřejněna krystalová struktura lidské Mad1 v komplexu s lidskou Mad2 tvořící tetramer. Kvůli experimentálním omezením struktura zobrazuje pouze zbytky Mad1 484 - 584. Prodloužené monomery Mad1 jsou pevně drženy pohromadě paralelní stočenou cívkou zahrnující N-koncové alfa šroubovice. Řetězy Mad1 směřují od stočené cívky směrem k jejich ligandům Mad2 a tvoří s Mad2 dva subkomplexy. Segment mezi alfa helixy 1 a 2 obsahuje vazebnou doménu Mad2. První část této vazebné domény je flexibilní a přijímá různé konformace, které vedou k asymetrickému komplexu. Ve své práci využívající termodynamické studie Sironi et al.[2]ukazují, že funkce Mad1 zpomaluje rychlost Mad2-CDC20 tvorba komplexu, a proto působí jako kompetitivní inhibitor in vivo. Autoři dále navrhují, že vazebná místa Mad1-Mad2 jsou pohřbena uvnitř struktury, což pravděpodobně činí vazebná místa nepřístupnými pro vazbu Cdc20. Vazba Mad1-Mad2 je neobvyklá v tom, že C-terminál Mad2 se přehýbá přes Mad1. Autoři proto dospěli k závěru, že nerušený komplex Mad1-Mad2 neuvolní Mad2 vyžadující nový, dosud špatně pochopený mechanismus konformačních změn.[2]
Rakovina
Nesrovnalosti v počtu chromozomů (aneuploidie) během meiózy jsou zodpovědné za lidská onemocnění, jako je Downov syndrom, a také se často objevují v rakovinných buňkách. Základní funkce SAC vede k hypotéze, že mutace SAC a zejména inaktivace SAC mohou být důvodem pro tumorigenezi nebo alespoň usnadnit tumorigenezi.[3] Proti této myšlence se ukázalo, že rakovinné buňky podstupují apoptózu, když nejsou přítomny složky SAC.[7] V tomto modelu se na rozdíl od jiného modelu stává inaktivace SAC potenciálním způsobem, jak zabít rychle se dělící rakovinné buňky. Molekulární vazby mezi Mad1p, SAC, apoptózou a rakovinou stále nejsou plně pochopeny.[3]
Viz také
Reference
- ^ A b C Hardwick KG, Murray AW (1995). „Mad1p, fosfoproteinová složka kontrolního bodu sestavy vřetena v nadějných kvasnicích“. The Journal of Cell Biology. 131 (3): 709–720. doi:10.1083 / jcb.131.3.709. PMC 2120625. PMID 7593191.
- ^ A b C Sironi L, Mapelli M, Knapp S, De Antoni A, Jeang KT, Musacchio A (2002). „Krystalová struktura tetramerického komplexu jádra Mad1 – Mad2: důsledky vazebného mechanismu„ bezpečnostního pásu “pro kontrolní bod vřetena“. Časopis EMBO. 21 (10): 2496–2506. doi:10.1093 / emboj / 21.10.2496. PMC 126000. PMID 12006501.
- ^ A b C d E F Musacchio A, Salmon ED (květen 2007). "Kontrolní bod sestavy vřetena v prostoru a čase". Nat. Rev. Mol. Cell Biol. 8 (5): 379–93. doi:10.1038 / nrm2163. PMID 17426725. S2CID 205494124.
- ^ Yu H (duben 2006). „Strukturální aktivace Mad2 v kontrolním bodě mitotického vřetena: dvoustavový model Mad2 versus model šablony Mad2“. J. Cell Biol. 173 (2): 153–157. doi:10.1083 / jcb.200601172. PMC 2063805. PMID 16636141.
- ^ Bharadwaj R, Yu H (2000). „Vřetenový kontrolní bod, aneuploidie a rakovina“. Onkogen. 23 (11): 2016–27. doi:10.1038 / sj.onc.1207374. PMID 15021889.
- ^ Chen RH, Shevchenko A, Mann M, Murray AW (1998). „Protein Xmad1 vřetenového kontrolního bodu získává Xmad2 do nepřipojených kinetochorů“. The Journal of Cell Biology. 143 (2): 283–295. doi:10.1083 / jcb.143.2.283. PMC 2132829. PMID 9786942.
- ^ Kops GJ, Foltz DR, Cleveland DW (červen 2004). „Letalita k lidským rakovinným buňkám prostřednictvím masivní ztráty chromozomů inhibicí mitotického kontrolního bodu“. Proc. Natl. Acad. Sci. USA. 101 (23): 8699–704. Bibcode:2004PNAS..101,8699 tis. doi:10.1073 / pnas.0401142101. PMC 423258. PMID 15159543.