Barevný střed - Colour centre - Wikipedia
| Barevný střed | |
|---|---|
 Oblast barevného vidění zobrazená jako V8 na horním obrázku | |
| Anatomická terminologie |
The barevný střed je oblast v mozku primárně zodpovědná za Vizuální vnímání a kortikální zpracování barevných signálů přijímaných okem, což nakonec vede k barevné vidění. Předpokládá se, že barevný střed u lidí je umístěn ve ventrální oblasti týlní lalok jako součást vizuální systém, kromě dalších oblastí odpovědných za rozpoznávání a zpracování konkrétního vizuálu podněty, například tváře, slova a objekty. Mnoho funkční zobrazování magnetickou rezonancí (fMRI) studie u lidí i makak opice prokázaly barevné podněty k aktivaci více oblastí v mozku, včetně fusiform gyrus a lingvální gyrus. Tyto oblasti, stejně jako další identifikované jako oblasti, které hrají roli ve zpracování barevného vidění, jsou společně označeny vizuální oblast 4 (V4). Přesné mechanismy, umístění a funkce V4 se stále zkoumají.
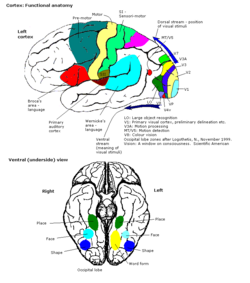
Primární zraková kůra
Primární část vizuální kůra, (V1), se nachází v kalkarinový sulcus, a je první kortikální oblast podílející se na vizuálním zpracování. Přijímá vizuální vstup z boční geniculate jádro, který se nachází v thalamus. V1 odesílá vizuální informace přijaté z LGN do jiných extrastriate kůra oblasti pro zpracování vyšších objednávek. Toto zpracování vyššího řádu zahrnuje rozpoznávání tvarů, pohybu a barev.[1]
Verze V1 má několik oblastí, které jsou citlivé na barvy, což znamená, že zpracování barev není omezeno na jednu oblast. Podle článku Dr. Roberta Shapleye má V1 důležitou roli ve vnímání barev. Experimentální výsledky fMRI ukázaly, že V1 má dva druhy neuronů citlivých na barvu: buňky s jedním soupeřem a buňky s dvojitým soupeřem. Tyto buňky jsou nedílnou součástí oponentský proces interpretace barevných signálů. Neurony s jedním soupeřem reagují na velké barevné oblasti. To je výhodné pro rozpoznávání velkých barevných scén a atmosfér. Pro srovnání, buňky dvojitého soupeře reagují na vzory, textury a hranice barev. To je důležitější pro vnímání barvy objektů a obrázků. Buňky s dvojitým protivníkem jsou vnímavé k opačným vstupům od různých kuželové buňky v sítnice. To je ideální pro identifikaci kontrastních barev, jako je červená a zelená. [1] Buňky s dvojitým protivníkem jsou zvláště důležité při výpočtu poměrů místních kuželů z vizuálních informací z nich receptivní pole.[1][2]
Jediné oponenty barevně citlivé neurony lze rozdělit do dvou kategorií v závislosti na signálech, které dostávají z buněk kužele: L-M neurony a S / (L + M) neurony. Tři typy kuželových buněk, malé (S), střední (M) a dlouhé (L), detekují různé vlnové délky napříč viditelné spektrum. Buňky kužele S mohou vidět barvy krátké vlnové délky, což odpovídá fialové a modré barvě. Podobně M buňky detekují barvy střední vlnové délky, jako je zelená a žlutá, a L buňky detekují barvy dlouhé vlnové délky, jako je červená. Neurony LM, nazývané také červeno-zelené protivníkovy buňky, přijímají vstup z kuželů s dlouhou vlnovou délkou, na rozdíl od vstupu z kuželů se střední vlnovou délkou. S / (L + M) neurony přijímají vstup z S-buněk a je proti součtu vstupů L a M-buněk. S / (L + M) neurony se také nazývají modro-žluté oponentní buňky. Protiklad mezi barvami umožňuje vizuálnímu systému interpretovat rozdíly v barvě, což je nakonec efektivnější než samostatné zpracování barev.[1][3]
Vizuální zpracování vyššího řádu
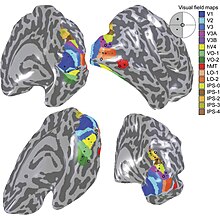
Primární vizuální kůra V1 odesílá vizuální informace do extrastriate kortikálních oblastí pro vizuální zpracování vyššího řádu. Tyto extrastriate kortikální oblasti jsou umístěny před týlním lalokem. Hlavní jsou označeny jako vizuální oblasti V2, V3, V4 a V5 / MT. Každá oblast může mít více funkcí. Nedávné nálezy ukázaly, že barevný střed není izolovaný ani sledovatelný k jediné oblasti ve zrakové kůře. Spíše existuje několik oblastí, které mohou mít různé role ve schopnosti zpracovávat barevný stimul.
Vizuální oblast V4
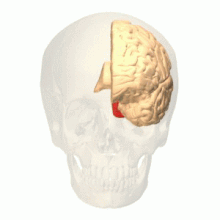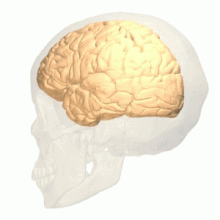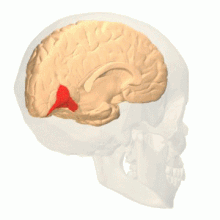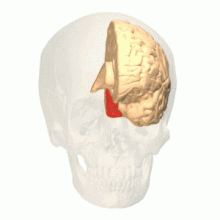

Anatomické a fyziologické studie prokázaly, že centrum barev začíná ve V1 a vysílá signály do extrastrovaných oblastí V2 a V4 pro další zpracování. Zejména V4 je oblastí zájmu kvůli síle barevných polí vnímajících jeho neurony.[4] V4 byla původně identifikována v pokusech s mozkovou kůrou makaků. Původně bylo navrženo, aby byla barva selektivně zpracována ve V4. Tato hypotéza však byla později odmítnuta ve prospěch jiné hypotézy, která naznačovala, že V4 a další oblasti kolem V4 spolupracují na zpracování barvy ve formě více barevných selektivních oblastí.[5] Po identifikaci V4 jako oblasti selektivní barvy u opic makaků začali vědci hledat homologní strukturu v lidské kůře. Pomocí fMRI zobrazování mozku vědci našli tři hlavní oblasti stimulované barvou: V1, oblast ventrálního týlního laloku, konkrétně lingvální gyrus, který byl označen jako lidský V4, nebo hV4, a další oblast umístěná vpředu ve fusiformním gyrus, označená jako jako V4α.[4][6]
Účel V4 se dynamicky měnil s prováděním nových studií. Vzhledem k tomu, že V4 silně reaguje na barvu u makaků i lidí, stala se oblastí zájmu vědců.[6] Oblast V4 byla původně přičítána selektivitě barev, ale nové důkazy ukázaly, že V4, stejně jako další oblasti zrakové kůry, jsou vnímavé k různým vstupům. Neurony V4 jsou vnímavé k řadě vlastností, jako je barva, jas a struktura. Rovněž se podílí na zpracování tvaru, orientace, zakřivení, pohybu a hloubky.[7]
Skutečná organizace hV4 v kůře se stále zkoumá. V opici makaků překlenuje V4 hřbetní a břišní týlní lalok. Lidské experimenty ukázaly, že V4 překlenuje pouze ventrální část. To vedlo k rozlišení hV4 od makaka V4. Nedávná studie Winawera a kol. analýza měření fMRI k mapování hV4 a ventrálních okcipitálních oblastí ukázala, že rozdíly mezi subjekty použitými pro mapování hV4 byly zpočátku připisovány chybě instrumentace, ale Winawer tvrdil, že dutiny v mozku interferovaly s měřením fMRI. Byly testovány dva modely pro hV4: jeden model měl hV4 úplně na břišní straně a druhý model měl hV4 rozdělen na hřbetní a břišní část. Byl vyvozen závěr, že je stále obtížné zmapovat aktivitu hV4 a že je nutné další vyšetřování. Existují však i další důkazy, jako jsou léze v ventrálním týlním laloku achromatopsie, navrhl, že ventrální okcipitální oblast hraje důležitou roli v barevném vidění.[8]
V4α
Hledání lidského ekvivalentu V4 vedlo k objevení dalších oblastí stimulovaných barvou. Nejvýznamnější byla oblast vpředu ve ventrálním týlním laloku, později pojmenovaná V4α. Další experimenty fMRI zjistily, že V4α má jinou funkci než V4, ale spolupracovaly s ní.[1] V4α je zapojen do řady procesů a je aktivní během úkolů vyžadujících řazení barev, snímky, znalosti o barvě, barevné iluze a barvu objektu.
Komplex V4-V4α
Oblasti V4 a V4α jsou samostatné entity, ale kvůli jejich těsné blízkosti ve fusiformním gyrusu jsou tyto dvě oblasti často souhrnně nazývány komplexem V4. Výzkum komplexu V4 zjistil, že různé chromatické stimulace aktivovaly buď oblast V4 nebo V4α, a některé parametry stimulace aktivovaly obě. Například přirozeně zbarvené obrázky aktivovaly V4α účinněji než V4. Nepřirozeně barevné obrázky aktivovaly V4α i V4 stejně. Byl učiněn závěr, že obě podskupiny spolupracují navzájem za účelem generování barevných obrazů, ale jsou také funkčně oddělené.[4]
Studie Nunn et al. o aktivaci komplexu V4 u lidí s vizuální synaesthesia ze slyšení mluvených slov bylo použito k předpovědi umístění centra barev. Synaesthesia je jev, kdy smyslový stimul produkuje automatickou a nedobrovolnou reakci u jiného pocit. V této studii byli studováni lidé, kteří by na sluchu viděli barvy, aby zjistili, zda lze barevnou reakci vysledovat do konkrétní kortikální oblasti. Výsledky fMRI ukázaly, že levý fusiformní gyrus, oblast shodná s V4, byl aktivován, když subjekty mluvily. Zjistili také současnou aktivaci V4α. V oblastech V1 a V2 byla malá aktivita. Tyto výsledky potvrdily existenci komplexu V4 u lidí jako oblasti specializované na barevné vidění.[9]
V2 prestriate cortex
Předpokládá se, že V2, nazývaná také prestriátová kůra, má malou roli ve zpracování barev promítáním signálů z V1 do komplexu V4. To, zda jsou ve V2 přítomny barevné selektivní buňky, se stále zkoumá. Některé studie optického zobrazování zjistily malé shluky červeno-zelených barevných selektivních buněk ve V1 a V2, ale ne žádné modro-žluté barevné selektivní buňky.[1] Další studie ukázaly, že V2 je aktivován barevnými podněty, ale ne barvou po obrázcích. [8] V4 má také zpětnou vazbu k V2, což naznačuje, že existuje definovaná síť komunikace mezi více oblastmi zrakové kůry. Když GABA, inhibiční neurotransmiter, byl injikován do buněk V4, došlo u buněk V2 k výraznému snížení excitability.[10]
Metody výzkumu
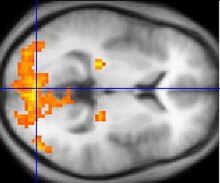
Funkční zobrazování magnetickou rezonancí, nebo zkráceně fMRI, byl klíčem při určování barevně selektivních oblastí ve vizuální kůře. fMRI je schopen sledovat mozkovou aktivitu měřením průtoku krve mozkem. Oblasti, které k nim proudí více krve, naznačují výskyt neuronální aktivity. Tato změna průtoku krve se nazývá hemodynamická odezva. Mezi výhody fMRI patří dynamické mapování kortikálních procesů v reálném čase. FMRI však nemůže sledovat skutečné pálení neuronů, ke kterému dochází v milisekundovém časovém měřítku, ale může sledovat hemodynamickou odezvu, která se děje v sekundovém časovém měřítku. Tato metoda je ideální pro sledování barevně selektivních neuronů, protože vnímání barev vede k vizuálu dosvit které lze pozorovat v neuronech, což trvá asi 15 sekund.[11]
Sakai a kol. použil fMRI ke sledování, zda aktivace fusiformního gyru korelovala s vnímáním barvy a následným obrazem. Subjekty ve studii Sakai byly umístěny do stroje fMRI a následně byly podrobeny různým vizuálním stimulům. Série tří obrazů byla ukázána subjektům, zatímco fMRI byl použit k zaměření na hemodynamiku fusiformního gyrusu. První obrázek byl vzor šesti barevných kruhů. Další dva obrázky byly achromatické. Jeden z obrazů měl šedý kříž a druhý obraz měl stejných šest kruhů jako první obraz, kromě toho, že šlo o šest odstínů šedé, které korelovaly s barevnými obrazy. Subjekty byly cyklovány mezi kruhovými a křížovými obrazy. Během křížových obrazů vnímal subjekt následný obraz. Výsledky experimentu ukázaly, že při pohledu na barevný obraz došlo k výraznému zvýšení aktivity fusiformního gyrusu. To poskytlo více důkazů o existenci barevného centra mimo primární vizuální kůru.[11]
Mozková achromatopsie
Mozková achromatopsie je chronický stav, kdy člověk není schopen vidět barvu, ale stále je schopen rozpoznat tvar a tvar. Mozková achromatopsie se liší od vrozené achromatopsie v tom, že je to způsobeno poškozením mozkové kůry na rozdíl od abnormalit v buňkách sítnice. Hledání barevného centra bylo motivováno objevem, že léze v ventrálním týlním laloku vedly k barvosleposti, stejně jako myšlenkou, že v mozkové kůře existují oblastní specializace. Mnoho studií ukázalo, že léze v oblastech běžně identifikovaných jako barevné centrum, jako jsou V1, V2 a komplex V4, vedou k achromatopsii.[1] Mozková achromatopsie nastává po poranění gyrusu lingválního nebo fusiformního, oblastí spojených s hV4. Tato zranění zahrnují fyzické trauma, mrtvici a růst nádoru. Jednou z hlavních iniciativ zaměřených na lokalizaci barevného centra ve zrakové kůře je odhalení příčiny a možné léčby mozkové achromatopsie.

Rozsah příznaků a poškození se u jednotlivých lidí liší. Pokud má člověk úplnou achromatopsii, nemá celé zorné pole zbarvené. Osoba s dyschromatopsií nebo neúplnou achromtopsií má podobné příznaky jako úplná achromatopsie, ale v menší míře. K tomu může dojít u lidí, kteří měli achromatopsii, ale mozek se zotavil ze zranění a obnovil určité barevné vidění. Osoba může vidět určité barvy. Existuje však mnoho případů, kdy nedojde k zotavení. Nakonec člověk s hemiachromatopsií vidí polovinu svého zorného pole barevně a druhou polovinu šedě. Vizuální hemifield kontralaterální k lézi v lingválním nebo fusiformním gyrusu je ten, který se jeví šedý, zatímco ipsilaterální vizuální hemifield má barvu.[11] Rozptyl symptomů zdůrazňuje potřebu porozumět architektuře barevného centra, aby bylo možné lépe diagnostikovat a léčit mozkovou achromotopsii.
Reference
- ^ A b C d E F Shapley R., Hawken M. J. (2011). „Color in the Cortex: single- and double-opell cells“. Vision Vision. 51 (7): 701–717. doi:10.1016 / j.visres.2011.02.012. PMC 3121536. PMID 21333672.
- ^ Conway BR (15. dubna 2001). "Prostorová struktura vstupů kužele do barevných buněk ve výstražné makakové primární vizuální kůře (V-1)". J. Neurosci. 21 (8): 2768–83. doi:10.1523 / JNEUROSCI.21-08-02768.2001. PMC 6762533. PMID 11306629.
- ^ Livingstone M. S., Hubel D. H. (1984). "Anatomie a fyziologie barevného systému ve zrakové kůře primátů". Journal of Neuroscience. 4: 309–356. doi:10.1523 / jneurosci.04-01-00309.1984. PMID 6198495.
- ^ A b C Bartels A., Zeki S. (2000). "Architektura barevného centra v lidském zrakovém mozku: nové výsledky a recenze". Evropský žurnál neurovědy. 12 (1): 172–193. doi:10.1046 / j.1460-9568.2000.00905.x. PMID 10651872.
- ^ Tootell R. B. H., Nelissen K., Vanduffel W., Orban G. A. (2004). „Hledat barevné„ středy “v Macaque Visual Cortex“. Mozková kůra. 14 (4): 353–363. doi:10.1093 / cercor / bhh001. PMID 15028640.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ A b Murphey D. K., Yoshor D., Beauchamp Michael S. (2008). "Vnímání odpovídá selektivitě ve středu lidské přední barvy". Aktuální biologie. 18 (3): 216–220. doi:10.1016 / j.cub.2008.01.013. PMID 18258428.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Roe Anna W. (2012). „Směrem k jednotné teorii vizuální oblasti V4“. Neuron. 74 (1): 12–29. doi:10.1016 / j.neuron.2012.03.011.
- ^ Winawer J., Horiguchi H., Sayres R. A., Amano K., Wandell B. A. (2010). „Mapování hV4 a ventrální týlní kůry: Zatmění žíly“. Journal of Vision. 10: 5. doi:10.1167/10.5.1.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Nunn J. A., Gregory L. J., Brammer M., Williams S. C. R., Parslow D. M., Morgan M. J., Gray J. A. (2002). "Funkční magnetická rezonance synestézie: aktivace V4 / V8 mluvenými slovy. [Článek]". Přírodní neurovědy. 5 (4): 371–375. doi:10.1038 / nn818. PMID 11914723.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Jansen-Amorim A. K., Fiorani M., Gattass R. (2012). „GABA inaktivace oblasti V4 mění vlastnosti receptivního pole neuronů V2 u opic Cebus“. Experimentální neurologie. 235 (2): 553–562. doi:10.1016 / j.expneurol.2012.03.008.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ A b C Sakai K., Watanabe E., Onodera Y., Uchida I., Kato H., Yamamoto E., Miyashita Y. (1995). „Funkční mapování lidského barevného centra pomocí echo-planární magnetické rezonance“. Sborník: Biologické vědy. 261 (1360): 89–98. doi:10.1098 / rspb.1995.0121.CS1 maint: více jmen: seznam autorů (odkaz)