Channelrhodopsin - Channelrhodopsin - Wikipedia
Channelrhodopsins jsou podrodinou retinylidenové proteiny (rhodopsiny ), které fungují jako světelné iontové kanály.[1] Slouží jako smyslové fotoreceptory jednobuněčné zelené řasy, ovládání fototaxe: pohyb v reakci na světlo.[2] Jsou vyjádřeny v buňkách jiných organismů a umožňují ovládání světla elektrická excitabilita, intracelulární kyselost, vápník příliv a další buněčné procesy (viz optogenetika ). Channelrhodopsin-1 (ChR1) a Channelrhodopsin-2 (ChR2) z modelového organismu Chlamydomonas reinhardtii jsou první objevené channelrhodopsiny. Varianty byly klonovány z jiných druhů řas a očekává se další.
Struktura
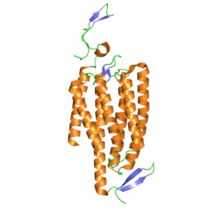
Pokud jde o strukturu, channelrhodopsiny jsou retinylidenové proteiny. Oni jsou sedm transmembránových proteinů jako rhodopsin a obsahují izomerizovatelné světlo chromofor Všechno-trans -sítnice (an aldehyd derivát z vitamin A. ). Chromofor sítnice je kovalentně spojen se zbytkem proteinu prostřednictvím protonace Schiffova základna. Zatímco většina 7-transmembránových proteinů je Receptory spojené s G proteinem které otevřou jiné iontové kanály nepřímo přes druhí poslové (tj. jsou metabotropní ), channelrhodopsiny přímo tvoří iontové kanály (tj. jsou ionotropní ).[4] Díky tomu je buněčná depolarizace extrémně rychlá, robustní a užitečná pro aplikace v oblasti bioinženýrství a neurovědy fotostimulace.
Funkce

Přírodní („divoký“ typ) ChR2 absorbuje modré světlo s absorpcí a akční spektrum maximálně při 480 nm.[5] Když všetrans-retinální komplex absorbuje a foton, vyvolává konformační změnu ze všehotrans do 13-cis-Retinal. Tato změna zavádí další v transmembránovém proteinu a otevírá póry alespoň na 6 Á. Během milisekund se sítnice uvolní zpět do formy all-trans, uzavře póry a zastaví tok iontů.[4] Většina přirozených channelrhodopsinů je nespecifických kation kanály, vedení H+, Na+, K.+a Ca2+ ionty. Nedávno, aniontově vodivé kanály rodopsiny byly objeveny.[6]
Designer-channelrhodopsins
Channelrhodopsins jsou klíčové nástroje optogenetika. The C-terminál konec Channelrhodopsinu-2 zasahuje do intracelulárního prostoru a může být nahrazen fluorescenční proteiny bez ovlivnění funkce kanálu. Tento druh fúzní konstrukt může být užitečné k vizualizaci morfologie buněk exprimujících ChR2.[7][8] Bodové mutace Bylo prokázáno, že blízko vazebné kapsy sítnice ovlivňují biofyzikální vlastnosti channelrhodopsinu, což vede k řadě různých nástrojů.
Kinetika
Uzavření kanálu po optické aktivaci lze podstatně oddálit mutací proteinových zbytků C128 nebo D156. Výsledkem této modifikace jsou supercitlivé kanálové rodopsiny, které lze otevřít pulzem modrého světla a uzavřít pulzem zeleného nebo žlutého světla (opční kroky s funkcí).[9][10][11] Mutace zbytku E123 urychluje kinetiku kanálu (ChETA) a výsledné mutanty ChR2 byly použity ke špičce neuronů až do 200 Hz.[12] Obecně platí, že channelrhodopsiny s pomalou kinetikou jsou na úrovni populace citlivější na světlo, protože otevřené kanály se časem hromadí i při nízké úrovni osvětlení.
Amplituda fotoproudu
Mutanty H134R a T159C vykazují zvýšené fotoproudy a kombinace T159 a E123 (ET / TC) má o něco větší fotoproudy a mírně rychlejší kinetiku než divoký typ ChR2.[13] Mezi variantami ChR vykazuje ChIEF, chiméra a bodový mutant ChR1 a ChR2 největší fotoproudy a nejméně desenzibilizaci a má kinetiku podobnou divokému typu ChR2.[14]
Vlnová délka
Chimérické kanálrhodopsiny byly vyvinuty kombinací transmembránových šroubovic z ChR1 a VChR1, což vedlo k vývoji ChR s červenými spektrálními posuny (jako C1V1 a ReaChR).[11][15] ReaChR zlepšil transport membrány a silnou expresi v savčích buňkách a byl použit pro minimálně invazivní transkraniální aktivaci mozkového kmene motoneurony. Hledání homologních sekvencí v jiných organismech přineslo spektrálně vylepšené a silnější červeně posunuté kanálové rodpsiny (Chrimson).[16] V kombinaci s ChR2 umožňují tyto žluté / červené světlocitlivé kanálové rodopsiny ovládat dvě populace neuronů nezávisle pomocí světelných pulsů různých barev.[17]
V řase byl objeven modře posunutý kanál rhodopsin Scherffelia dubia. Po nějakém inženýrství pro zlepšení přenosu a rychlosti membrány výsledný nástroj (CheRiff) vytvořil velké fotoproudy při excitaci 460 nm.[18] Byl kombinován s geneticky kódovaným indikátorem vápníku jRCaMP1b [19] v plně optickém systému zvaném OptoCaMP.[20]
Iontová selektivita
Mutace L132C (CatCh) zvyšuje propustnost pro vápník a generuje velmi velké proudy.[21] Mutace E90 na kladně nabitou aminokyselinu arginin přemění channelrhodopsin z kanálu nespecifického kationu na kanál vedoucí chlorid (ChloC).[22] Selektivita pro Cl- byla dále zlepšena nahrazením záporně nabitých zbytků v póru kanálu, čímž se vytvořila reverzní potenciál negativnější.[23][24] Selektivní chlorid vodivý kanál rhodopsiny (iChloC, iC ++, GtACR) inhibují růst neuronů v buněčné kultuře a u intaktních zvířat, když jsou osvětleny modrým světlem.
Aplikace
Channelrhodopsiny lze snadno exprimovat v excitovatelných buňkách, jako je neurony pomocí různých transfekčních technik (virové transfekce, elektroporace, genová zbraň ) nebo transgenní zvířata. Světlo absorbující pigment sítnice je přítomen ve většině buněk (z obratlovců ) tak jako Vitamin A, což umožňuje fotostimulovat neurony bez přidání jakýchkoli chemických sloučenin. Před objevem channelrhodopsinů byli neurologové omezeni na záznam aktivita neuronů v mozku a korelát tuto činnost s chováním. To nestačí k prokázání, že zaznamenaná nervová aktivita ve skutečnosti způsobil toto chování. Ovládání sítí geneticky modifikovaných buněk světlem, vznikající pole známé jako Optogenetika., umožňuje vědcům nyní prozkoumat kauzální souvislost mezi aktivitou ve specifické skupině neuronů a duševní události, např. rozhodování. Optická kontrola chování byla prokázána u hlístic, ovocných mušek, zebrafish a myší.[25][26] Nedávno, chlorid vodivý kanál rhodopsiny byly zkonstruovány a byly také nalezeny v přírodě.[6][22] Tyto nástroje lze použít k umlčení neuronů v buněčné kultuře a v živých zvířatech pomocí inhibice posunu.[23][24]
Použití více barev světla rozšiřuje možnosti optogenetický experimenty. ChR2 citlivý na modré světlo a čerpadlo chloridu aktivovaného žlutým světlem halorhodopsin společně umožňují vícebarevnou optickou aktivaci a umlčení nervové aktivity.[27][28] VChR1 z koloniální řasy Volvox carteri absorbuje maximálně při 535 nm a byl použit ke stimulaci buněk žlutým světlem (580 nm), i když světelné proudy generované VChR1 jsou obvykle velmi malé.[29] Hybridy VChR1-ChR2 však byly vyvinuty pomocí řízená evoluce které zobrazují maximální buzení při 560 nm a 50% špičkové absorbance při vlnových délkách nad 600 nm.[15][30]
Pomocí fluorescenčně značeného ChR2, stimulovaného světlem axony a synapse lze identifikovat.[8] To je užitečné ke studiu molekulárních událostí během indukce synaptická plasticita.[31] Transfektovaný kultivované neuronové sítě lze stimulovat k provádění požadovaného chování pro aplikace v robotice a řízení.[32] ChR2 se také používá k mapování spojení na velké vzdálenosti z jedné strany mozku na druhou a k mapování prostorového umístění vstupů na dendritickém stromu jednotlivých neuronů.[33][34]
Vizuální funkce u slepých myší může být částečně obnovena expresí ChR2 ve buňkách vnitřní sítnice.[35][36] V budoucnu může ChR2 najít lékařské aplikace, např. ve formách degenerace sítnice nebo pro hluboká mozková stimulace. Optický kochleární implantáty Bylo prokázáno, že dobře fungují při pokusech na zvířatech a mohly by vést k první aplikaci optogenetika u lidských pacientů.[37][38][39]
Dějiny
Pohyblivost a fotoorientace mikrořas (fototaxe ) byly studovány více než sto let v mnoha laboratořích po celém světě.
V roce 1980 vyvinul Ken Foster první konzistentní teorii o funkčnosti řasových očí.[40] Analyzoval také publikovaná akční spektra a doplnil slepé buňky retinálními a retinálními analogy, což vedlo k závěru, že fotoreceptor pro odpovědi motility v Chlorophyceae je rhodopsin.[41]
Fotoproudy Chlorophyceae Heamatococcus pluvialis a Chlamydomonas reinhardtii byly po mnoho let studovány ve skupinách Olega Sineshchekova a Peter Hegemann, jehož výsledkem byly dvě klíčové publikace v letech 1978 a 1991.[42][43] Na základě akční spektroskopie a simultánních záznamů fotoproudů a bičíkových výbojů bylo stanoveno, že fotoreceptorové proudy a následné bičíkové pohyby jsou zprostředkovány rhodopsinem a kontrolní fototaxí a fotofobními reakcemi. Extrémně rychlý vzestup fotoreceptorového proudu po krátkém světelném záblesku vedl k závěru, že rhodopsin a kanál jsou úzce spojeny v proteinovém komplexu nebo dokonce v jednom jediném proteinu.[44][45]
Biochemická purifikace rhodopsin-fotoreceptoru (ů) však byla po mnoho let neúspěšná.
Nukleotidové sekvence rhodopsinů, nyní nazývaných channelrhodopsins ChR1 a ChR2, byly nakonec odkryty ve velkém měřítku EST sekvenční projekt v C. reinhardtii. Nezávislé předkládání stejných sekvencí do GenBank tři výzkumné skupiny vyvolaly zmatek ohledně jejich pojmenování: Jména policajt-3 a policajt-4 byly použity pro počáteční podání Hegemannovou skupinou;[46] csoA a csoB Spudichova skupina;[2] a acop-1 a acop-2 skupinou Takahashi.[47] Bylo zjištěno, že obě sekvence fungují jako jednosložkové kationtové kanály aktivované světlem Xenopus oocyty a buňky lidských ledvin (HEK) od Georga Nagela, Ernsta Bamberga, Petera Hegemanna a dalších.[1][4]
Název „channelrhodopsin“ byl vytvořen pro zvýraznění této neobvyklé vlastnosti a sekvence byly odpovídajícím způsobem přejmenovány. Mezitím jejich role při generování fotoreceptorových proudů v buňkách řas charakterizovali Oleg Sineshchekov, Kwang-Hwan Jung a John Spudich,[2] a Peter Berthold a Peter Hegemann.[48]
V listopadu 2004 Zhuo-Hua Pan zaslal příspěvek do Příroda hlášení obnovy zraku u slepých myší transfektovaných Channelrhodopsinem,[Citace je zapotřebí ] ale papír byl odmítnut[Citace je zapotřebí ] a nakonec publikováno v Neuron v roce 2006.[Citace je zapotřebí ]
Mezitím, v roce 2005, tři skupiny postupně zavedly ChR2 jako nástroj pro geneticky cílené optické dálkové ovládání (optogenetika ) z neurony neurální obvody a chování.
Nejprve, Karl Deisseroth Laboratoř (v článku publikovaném v srpnu 2005) prokázala, že ChR2 lze nasadit k ovládání savčí neurony in vitro, dosažení časové přesnosti řádově v milisekundách (jak z hlediska zpoždění, tak i časového jitteru).[7] To bylo významné zjištění, protože zaprvé vyžadují všechny opsiny (mikrobiální i obratlovci) sítnice jako kofaktor pro snímání světla a nebylo jasné, zda centrální nervové buňky savců budou obsahovat dostatečné hladiny sítnice, ale obsahují; zadruhé, ukázalo se, navzdory malé jednokanálové vodivosti, dostatečnou účinnost k tomu, aby vedly savčí neurony nad prahovou hodnotu akčního potenciálu; a za třetí prokázal, že channelrhodopsin je prvním optogenetickým nástrojem, pomocí kterého by mohla být řízena nervová aktivita s časovou přesností, při které neurony fungují (milisekundy). Dřívější nástroj pro fotostimulaci, cHARGe, prokázal princip v kultivovaných neuronech[49] ale nikdy nebyl používán jinými skupinami, protože fungoval s přesností v řádu sekund, byl vysoce variabilní a neumožňoval ovládání jednotlivce akční potenciály.
Druhá studie byla zveřejněna později skupinami Petera Hegemanna a Stefana Herlitzeho, které potvrzují schopnost ChR2 řídit aktivitu obratlovců neurony, v této době v kuřecí míchě.[50] Tato studie byla první, kde byl ChR2 exprimován spolu s optickým tlumičem, obratlovcem rhodopsin -4 v tomto případě, demonstrace poprvé, že excitovatelné buňky mohly být aktivovány a umlčeny pomocí těchto dvou nástrojů současně, osvětlujících tkáň na různých vlnových délkách.
Skupiny Alexander Gottschalk a Ernst Bamberg (s experimentálním vedením Georga Nagela) prokázali, že ChR2, pokud je exprimován ve specifických neuronech nebo svalových buňkách, může vyvolat předvídatelné chování, tj. Může řídit nervový systém neporušeného zvířete, v tomto případě bezobratlý C. elegans.[51] Toto bylo první použití ChR2 k řízení chování zvířete v optogenetickém experimentu, kdy byl geneticky specifikovaný typ buňky podroben optickému dálkovému ovládání. Ačkoli oba aspekty byly dříve v tomto roce ilustrovány jinou skupinou, Miesenböck laboratoř využívající nepřímo světelný iontový kanál P2X2,[52] od nynějška dominovaly v oblasti geneticky cíleného dálkového ovládání excitabilních buněk mikrobiální opsiny jako channelrhodopsin, kvůli síle, rychlosti, cílitelnosti, snadnosti použití a časové přesnosti přímé optické aktivace, nevyžadující žádnou externí chemickou sloučeninu, například v kleci ligandy.[53]
Abychom překonali jeho hlavní nevýhody - malou jednokanálovou vodivost (zejména v ustáleném stavu), omezení na jednu optimální vlnovou délku buzení (~ 470 nm, modrá) a také relativně dlouhou dobu zotavení, která neumožňuje kontrolované odpálení neuronů výše 20–40 Hz - ChR2 byl optimalizován pomocí genetické inženýrství. A bodová mutace H134R (výměna aminokyseliny histidinu v poloze 134 nativního proteinu za arginin) vedla ke zvýšení vodivosti v ustáleném stavu, jak je popsáno v dokumentu z roku 2005, který také zavedl ChR2 jako optogenetický nástroj C. elegans.[51] V roce 2009, Roger Tsien Laboratoř optimalizovala ChR2 pro další zvýšení vodivosti v ustáleném stavu a dramaticky snížila desenzibilizaci vytvořením chimér ChR1 a ChR2 a mutací specifických aminokyselin, čímž se získaly ChEF a ChIEF, které umožnily řízení sledů akčních potenciálů až do 100 Hz.[14][54] V roce 2010 zavedly skupiny Hegemann a Deisseroth mutaci E123T do nativního ChR2, čímž se získala ChETA, která má rychlejší zapnutí a vypnutíkinetika, umožňující řízení jednotlivých akčních potenciálů při frekvencích do 200 Hz (u příslušných typů článků).[12][14]
Skupiny Hegemanna a Deisserotha také zjistily, že zavedení bodové mutace C128S činí z výsledného derivátu ChR2 nástroj krokové funkce: Jakmile je „zapnuto“ modrým světlem, zůstane ChR2 (C128S) v otevřeném stavu, dokud není přepnuto vypnuto žlutým světlem - modifikace, která zhoršuje časovou přesnost, ale zvyšuje citlivost na světlo o dva řády.[9] Objevili a charakterizovali také VChR1 v mnohobuněčných řasách Volvox carteri. VChR1 produkuje pouze malé fotoproudy, ale s absorpčním spektrem, které je oproti ChR2 posunuto červeně.[29] Použitím částí sekvence ChR1 byla později vylepšena fotoproudová amplituda, která umožnila excitaci dvou populací neuronů při dvou odlišných vlnových délkách.[11]
Deisserothova skupina je průkopníkem mnoha aplikací u živých zvířat, jako je například geneticky cílené dálkové ovládání hlodavci in vivo,[55] optogenetická indukce učení u hlodavců,[56] experimentální léčba Parkinsonova choroba u krys,[57][58] a kombinace s fMRI (opto-fMRI).[59] Jiné laboratoře propagovaly kombinaci stimulace ChR2 s zobrazováním vápníku pro všechny optické experimenty,[8] mapování dlouhého dosahu[33] a místní[60] neurální obvody, exprese ChR2 z transgenního lokusu - přímo[61] nebo v Cre-lox podmíněné paradigma[60] - stejně jako dva fotony excitace ChR2, umožňující aktivaci jednotlivých buněk.[62][63][64]
V březnu 2013 byla Brain Prize (Grete Lundbeck European Brain Research Prize) společně udělena Bambergovi, Boydenovi, Deisserothovi, Hegemannovi, Miesenböckovi a Nagelovi za „jejich vynález a zdokonalení optogenetiky“.[65] Ve stejném roce obdrželi Hegemann a Nagel Louis-Jeantetova cena za medicínu pro „objev channelrhodopsinu“. V roce 2015 obdrželi Boyden a Deisseroth Průlomová cena v biologických vědách a v roce 2020 obdrželi Miesenböck, Hegemann a Nagel Shaw cena v Life Science a medicíně pro vývoj optogenetiky.
Reference
- ^ A b Nagel G, Ollig D, Fuhrmann M, Kateriya S, Musti AM, Bamberg E, Hegemann P (červen 2002). „Channelrhodopsin-1: protonový kanál se zelenými bránami se zelenými řasami“. Věda. 296 (5577): 2395–8. doi:10.1126 / science.1072068. PMID 12089443.
- ^ A b C Sineshchekov OA, Jung KH, Spudich JL (červen 2002). „Dva rhodopsiny zprostředkovávají fototaxi na světlo s nízkou a vysokou intenzitou u Chlamydomonas reinhardtii“. Proc. Natl. Acad. Sci. USA. 99 (13): 8689–94. doi:10.1073 / pnas.122243399. PMC 124360. PMID 12060707.
- ^ Kato HE, Zhang F, Yizhar O, Ramakrishnan C, Nishizawa T, Hirata K, Ito J, Aita Y, Tsukazaki T, Hayashi S, Hegemann P, Maturana AD, Ishitani R, Deisseroth K, Nureki O (únor 2012). "Krystalová struktura kanálového kanálu rhodopsinu se světelnou bránou". Příroda. 482 (7385): 369–74. doi:10.1038 / příroda 10870. PMC 4160518. PMID 22266941.
- ^ A b C Nagel G, Szellas T, Huhn W, Kateriya S, Adeishvili N, Berthold P, Ollig D, Hegemann P, Bamberg E (listopad 2003). „Channelrhodopsin-2, přímo světelný kation-selektivní membránový kanál“. Proc. Natl. Acad. Sci. USA. 100 (24): 13940–5. doi:10.1073 / pnas.1936192100. PMC 283525. PMID 14615590.
- ^ Bamann C, Kirsch T, Nagel G, Bamberg E (leden 2008). "Spektrální charakteristiky fotocyklu channelrhodopsinu-2 a jeho důsledky pro funkci kanálu". J. Mol. Biol. 375 (3): 686–94. doi:10.1016 / j.jmb.2007.10.072. PMID 18037436.
- ^ A b Govorunova EG, Sineshchekov OA, Janz R, Liu X, Spudich JL (2015). „Aniontové kanály s přirozeným světlem: Rodina mikrobiálních rodopsinů pro pokročilou optogenetiku“. Věda. 349 (6248): 647–650. doi:10.1126 / science.aaa7484. PMC 4764398. PMID 26113638.
- ^ A b Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K (září 2005). "Milisekundový čas, geneticky cílená optická kontrola nervové aktivity". Nat. Neurosci. 8 (9): 1263–8. doi:10.1038 / nn1525. PMID 16116447.
- ^ A b C Zhang YP, Oertner TG (únor 2007). "Optická indukce synaptické plasticity pomocí kanálu citlivého na světlo". Nat. Metody. 4 (2): 139–41. doi:10.1038 / nmeth988. PMID 17195846.
- ^ A b Berndt A, Yizhar O, Gunaydin LA, Hegemann P, Deisseroth K (únor 2009). "Bistabilní neurální stavové přepínače". Nat. Neurosci. 12 (2): 229–34. doi:10.1038 / nn.2247. PMID 19079251.
- ^ Schoenenberger P, Gerosa D, Oertner TG (2009). „Časová kontrola okamžité časné indukce genu světlem“. PLOS ONE. 4 (12): e8185. doi:10,1371 / journal.pone.0008185. PMC 2780714. PMID 19997631.
- ^ A b C Yizhar O, Fenno LE, Prigge M, Schneider F, Davidson TJ, O'Shea DJ, Sohal VS, Goshen I, Finkelstein J, Paz JT, Stehfest K, Fudim R, Ramakrishnan C, Huguenard JR, Hegemann P, Deisseroth K ( Září 2011). „Neokortikální rovnováha excitace / inhibice při zpracování informací a sociální dysfunkce“. Příroda. 477 (7363): 171–8. doi:10.1038 / příroda10360. PMC 4155501. PMID 21796121.
- ^ A b Gunaydin LA, Yizhar O, Berndt A, Sohal VS, Deisseroth K, Hegemann P (březen 2010). "Ultrarychlá optogenetická kontrola". Nat. Neurosci. 13 (3): 387–92. doi:10.1038 / č. 2495. PMID 20081849.
- ^ Berndt A, Schoenenberger P, Mattis J, Tye KM, Deisseroth K, Hegemann P, Oertner TG (květen 2011). „Vysoce efektivní channelrhodopsiny pro rychlou neuronální stimulaci při slabém osvětlení“. Sborník Národní akademie věd Spojených států amerických. 108 (18): 7595–600. doi:10.1073 / pnas.1017210108. PMC 3088623. PMID 21504945.
- ^ A b C Lin JY (leden 2011). „Uživatelská příručka k variantám channelrhodopsinu: funkce, omezení a budoucí vývoj“. Experimentální fyziologie. 96 (1): 19–25. doi:10.1113 / expphysiol.2009.051961. PMC 2995811. PMID 20621963.
- ^ A b Lin JY, Knutsen PM, Muller A, Kleinfeld D, Tsien RY (říjen 2013). „ReaChR: červeně posunutá varianta channelrhodopsinu umožňuje hlubokou transkraniální optogenetickou excitaci“. Přírodní neurovědy. 16 (10): 1499–508. doi:10.1038 / č. 3502. PMC 3793847. PMID 23995068.
- ^ Klapoetke NC, Murata Y, Kim SS, Pulver SR, Birdsey-Benson A, Cho YK, Morimoto TK, Chuong AS, Carpenter EJ, Tian Z, Wang J, Xie Y, Yan Z, Zhang Y, Chow BY, Surek B, Melkonian M, Jayaraman V, Constantine-Paton M, Wong GK, Boyden ES (březen 2014). "Nezávislá optická excitace odlišných neurálních populací". Přírodní metody. 11 (3): 338–46. doi:10.1038 / nmeth.2836. PMC 3943671. PMID 24509633.
- ^ Háčky BM, Lin JY, Guo C, Svoboda K (březen 2015). „Dvoukanálové mapování obvodů odhaluje konvergenci senzomotorů v primární motorické kůře“. The Journal of Neuroscience. 35 (10): 4418–26. doi:10.1523 / JNEUROSCI.3741-14.2015. PMC 4355205. PMID 25762684.
- ^ Hochbaum DR, Zhao Y, Farhi SL, Klapoetke N, Werley CA, Kapoor V, Zou P, Kralj JM, Maclaurin D, Smedemark-Margulies N, Saulnier JL, Boulting GL, Straub C, Cho YK, Melkonian M, Wong GK, Harrison DJ, Murthy VN, Sabatini BL, Boyden ES, Campbell RE, Cohen AE (srpen 2014). "All-optická elektrofyziologie v savčích neuronech pomocí vytvořených mikrobiálních rodopsinů". Přírodní metody. 11 (8): 825–33. doi:10,1038 / nmeth.3000. PMC 4117813. PMID 24952910.
- ^ Dana H, Mohar B, Sun Y, Narayan S, Gordus A, Hasseman JP, Tsegaye G, Holt GT, Hu A, Walpita D, Patel R, Macklin JJ, Bargmann CI, Ahrens MB, Schreiter ER, Jayaraman V, Looger LL , Svoboda K, Kim DS (březen 2016). „Citlivé indikátory vápníku červené bílkoviny pro zobrazování nervové aktivity“. eLife. 5. doi:10,7554 / eLife.12727. PMC 4846379. PMID 27011354.
- ^ Afshar Sabre W, Gasparoli FM, Dirks MG, Gunn-Moore FJ, Antkowiak M (2018). „All-Optical Assay to Study Biological Neural Networks“. Frontiers in Neuroscience. 12: 451. doi:10.3389 / fnins.2018.00451. PMC 6041400. PMID 30026684.
- ^ Kleinlogel S, Feldbauer K, Dempski RE, Fotis H, Wood PG, Bamann C, Bamberg E (duben 2011). „Mimořádně citlivá na světlo a rychlá neuronální aktivace s Ca² + propustným kanálem rhodopsinem CatCh“ (PDF). Přírodní neurovědy. 14 (4): 513–8. doi:10.1038 / nn.2776. PMID 21399632.
- ^ A b Wietek J, Wiegert JS, Adeishvili N, Schneider F, Watanabe H, Tsunoda SP, Vogt A, Elstner M, Oertner TG, Hegemann P (duben 2014). "Konverze channelrhodopsinu na chloridový kanál se světelnou bránou". Věda. 344 (6182): 409–12. doi:10.1126 / science.1249375. PMID 24674867.
- ^ A b Wietek J, Beltramo R, Scanziani M, Hegemann P, Oertner TG, Wiegert JS (říjen 2015). "Vylepšený chlorodový kanálrhodopsin pro světlo indukovanou inhibici neuronální aktivity in vivo". Vědecké zprávy. 5: 14807. doi:10.1038 / srep14807. PMC 4595828. PMID 26443033.
- ^ A b Berndt A, Lee SY, Wietek J, Ramakrishnan C, Steinberg EE, Rashid AJ, Kim H, Park S, Santoro A, Frankland PW, Iyer SM, Pak S, Ährlund-Richter S, Delp SL, Malenka RC, Josselyn SA, Carlén M, Hegemann P, Deisseroth K (leden 2016). "Strukturální základy optogenetiky: Determinanty selektivity iontů kanálrhodopsinu". Sborník Národní akademie věd Spojených států amerických. 113 (4): 822–9. doi:10.1073 / pnas.1523341113. PMC 4743797. PMID 26699459.
- ^ Douglass AD, Kraves S, Deisseroth K, Schier AF, Engert F (srpen 2008). „Únikové chování vyvolané jedinými hroty vyvolanými kanálem rhodopsin-2 v somatosenzorických neuronech zebrafish“. Curr. Biol. 18 (15): 1133–7. doi:10.1016 / j.cub.2008.06.077. PMC 2891506. PMID 18682213.
- ^ Huber D, Petreanu L, Ghitani N, Ranade S, Hromádka T, Mainen Z, Svoboda K (leden 2008). „Řídká optická mikrostimulace v hlavové kůře pohání naučené chování u volně se pohybujících myší“. Příroda. 451 (7174): 61–4. doi:10.1038 / nature06445. PMC 3425380. PMID 18094685.
- ^ Han X, Boyden ES (2007). „Vícebarevná optická aktivace, umlčení a desynchronizace nervové aktivity s časovým rozlišením jednoho hrotu“. PLOS ONE. 2 (3): e299. doi:10.1371 / journal.pone.0000299. PMC 1808431. PMID 17375185.
- ^ Zhang F, Wang LP, Brauner M, Liewald JF, Kay K, Watzke N, Wood PG, Bamberg E, Nagel G, Gottschalk A, Deisseroth K (duben 2007). "Multimodální rychlý optický dotaz neurálních obvodů". Příroda. 446 (7136): 633–9. doi:10.1038 / nature05744. PMID 17410168.
- ^ A b Zhang F, Prigge M, Beyrière F, Tsunoda SP, Mattis J, Yizhar O, Hegemann P, Deisseroth K (červen 2008). „Optogenetická excitace s červeným posunem: nástroj pro rychlou neurální kontrolu odvozený od Volvox carteri“. Nat. Neurosci. 11 (6): 631–3. doi:10.1038 / č. 2120. PMC 2692303. PMID 18432196.
- ^ Yizhar O, Fenno LE, Prigge M, Schneider F, Davidson TJ, O'Shea DJ, Sohal VS, Goshen I, Finkelstein J, Paz JT, Stehfest K, Fudim R, Ramakrishnan C, Huguenard JR, Hegemann P, Deisseroth K ( Července 2011). „Neokortikální rovnováha excitace / inhibice při zpracování informací a sociální dysfunkce“. Příroda. 477 (7363): 171–8. doi:10.1038 / příroda10360. PMC 4155501. PMID 21796121.
- ^ Zhang YP, Holbro N, Oertner TG (srpen 2008). „Optická indukce plasticity v jednotlivých synapsích odhaluje akumulaci alfaCaMKII specifickou pro vstup“. Proc. Natl. Acad. Sci. USA. 105 (33): 12039–44. doi:10.1073 / pnas.0802940105. PMC 2575337. PMID 18697934.
- ^ Xu Z, Ziye X, Craig H, Silvia F (prosinec 2013). Špičkový nepřímý trénink virtuálního hmyzu řízeného neuronovou sítí. Rozhodování a řízení IEEE. 6798–6805. CiteSeerX 10.1.1.671.6351. doi:10.1109 / CDC.2013.6760966. ISBN 978-1-4673-5717-3.
- ^ A b Petreanu L, Huber D, Sobczyk A, Svoboda K (květen 2007). "Mapování obvodu s asistovaným kanálem rhodopsin-2 dalekonosných callosálních projekcí". Nat. Neurosci. 10 (5): 663–8. doi:10.1038 / nn1891. PMID 17435752.
- ^ Petreanu L, Mao T, Sternson SM, Svoboda K (únor 2009). „Subcelulární organizace neokortikálních excitačních spojení“. Příroda. 457 (7233): 1142–5. doi:10.1038 / nature07709. PMC 2745650. PMID 19151697.
- ^ Bi A, Cui J, Ma YP, Olshevskaya E, Pu M, Dizhoor AM, Pan ZH (duben 2006). „Ektopická exprese rhodopsinu mikrobiálního typu obnovuje vizuální odpovědi u myší s degenerací fotoreceptorů“. Neuron. 50 (1): 23–33. doi:10.1016 / j.neuron.2006.02.026. PMC 1459045. PMID 16600853.
- ^ Lagali PS, Balya D, Awatramani GB, Münch TA, Kim DS, Busskamp V, Cepko CL, Roska B (červen 2008). "Světlem aktivované kanály zaměřené na ON bipolární buňky obnovují vizuální funkci při degeneraci sítnice". Nat. Neurosci. 11 (6): 667–75. doi:10.1038 / nn.2117. PMID 18432197.
- ^ Hernandez VH, Gehrt A, Reuter K, Jing Z, Jeschke M, Mendoza Schulz A, Hoch G, Bartels M, Vogt G, Garnham CW, Yawo H, Fukazawa Y, Augustine GJ, Bamberg E, Kügler S, Salditt T, de Hoz L, Strenzke N, Moser T (únor 2014). „Optogenetická stimulace sluchové dráhy“. J Clin Invest. 124 (3): 1114–29. doi:10,1172 / JCI69050. PMC 3934189. PMID 24509078.
- ^ Mager T, Lopez de la Morena D, Senn V, Schlotte J, D'Errico A, Feldbauer K, Wrobel C, Jung S, Bodensiek K, Rankovic V, Browne L, Huet A, Jüttner J, Wood PG, Letzkus JJ, Moser T, Bamberg E (květen 2018). „Vysokofrekvenční neurální stouply a sluchová signalizace ultrarychlou červeně posunutou optogenetikou“. Nat Commun. 9 (1): 1750. doi:10.1038 / s41467-018-04146-3. PMC 5931537. PMID 29717130.
- ^ Keppeler D, Martins Merino R, Lopez de la Morena D, Bali B, Huet AT, Gehrt A, Wrobel C, Subramanian S, Dombrowski T, Wolf F, Rankovic V, Neef A, Moser T (2018). „Ultrarychlá optogenetická stimulace sluchové dráhy cíleně optimalizovaným Chronosem“. EMBO J.. 37 (24): e99649. doi:10.15252 / embj.201899649. PMC 6293277. PMID 30396994.
- ^ Foster KW, Smyth R (1980). „Světelné antény ve fototaktických řasách“. Mikrobiologické recenze. 44 (4): 572–630. PMC 373196. PMID 7010112.
- ^ Foster KW, Saranak J, Patel N, Zarilli G, Okabe M, Kline T, Nakanishi K (říjen 1984). „Rhodopsin je funkční fotoreceptor pro fototaxi v jednobuněčném eukaryotu Chlamydomonas.“ Příroda. 311 (5988): 489–491. doi:10.1038 / 311756a0. PMID 6493336.
- ^ Litvin FF, Sineshchekov OA, Sineshchekov VA (1978). "Fotoreceptorový elektrický potenciál ve fototaxi řasy Haematococcus pluvialis". Příroda. 271 (5644): 476–478. doi:10.1038 / 271476a0. PMID 628427.
- ^ Harz H, Hegemann P (červen 1991). "Rhodopsinem regulované proudy vápníku v Chlamydomonas". Příroda. 351 (6326): 489–491. doi:10.1038 / 351489a0.
- ^ Holland EM, Braun FJ, Nonnengässer C, Harz H, Hegemann P (únor 1996). "Povaha fotopřítoků spouštěných rhodopsinem v Chlamydomonas. I. Kinetika a vliv dvojmocných iontů". Biophys. J. 70 (2): 924–931. doi:10.1016 / S0006-3495 (96) 79635-2. PMC 1224992. PMID 8789109.
- ^ Braun FJ, Hegemann P (březen 1999). „Dvě vodivosti aktivované světlem v oku zelené řasy Volvox carteri“. Biophys. J. 76 (3): 1668–1778. doi:10.1016 / S0006-3495 (99) 77326-1. PMC 1300143. PMID 10049347.
- ^ Kateriya, S. Fuhrmann, M. Hegemann, P .: Přímé podání: gen pro retinální vazebný protein (cop4) pro Chlamydomonas reinhardtii; GenBank přístupové číslo AF461397
- ^ Suzuki T, Yamasaki K, Fujita S, Oda K, Iseki M, Yoshida K, Watanabe M, Daiyasu H, Toh H, Asamizu E, Tabata S, Miura K, Fukuzawa H, Nakamura S, Takahashi T (únor 2003). "Rhodopsiny archaeálního typu v Chlamydomonas: modelová struktura a intracelulární lokalizace". Biochem. Biophys. Res. Commun. 301 (3): 711–7. doi:10.1016 / S0006-291X (02) 03079-6. PMID 12565839.
- ^ Berthold P, Tsunoda SP, Ernst OP, Mages W, Gradmann D, Hegemann P (červen 2008). „Channelrhodopsin-1 iniciuje fototaxi a fotofobní reakce u chlamydomonas okamžitou depolarizací vyvolanou světlem“. Rostlinná buňka. 20 (6): 1665–1677. doi:10.1105 / tpc.108.057919. PMC 2483371. PMID 18552201.
- ^ Zemelman BV, Lee GA, Ng M, Miesenböck G (leden 2002). "Selektivní fotostimulace geneticky CHARGED neuronů". Neuron. 33 (1): 15–22. doi:10.1016 / S0896-6273 (01) 00574-8. PMID 11779476.
- ^ Li X, Gutierrez DV, Hanson MG, Han J, Mark MD, Chiel H, Hegemann P, Landmesser LT, Herlitze S (prosinec 2005). "Rychlá neinvazivní aktivace a inhibice nervové a síťové aktivity obratlovcem rhodopsinem a zelenými řasami channelrhodopsin". Proc. Natl. Acad. Sci. USA. 102 (49): 17816–21. doi:10.1073 / pnas.0509030102. PMC 1292990. PMID 16306259.
- ^ A b Nagel G, Brauner M, Liewald JF, Adeishvili N, Bamberg E, Gottschalk A (prosinec 2005). „Světelná aktivace channelrhodopsinu-2 v excitovatelných buňkách Caenorhabditis elegans spouští rychlé reakce v chování“. Curr. Biol. 15 (24): 2279–84. doi:10.1016 / j.cub.2005.11.032. PMID 16360690.
- ^ Lima SQ, Miesenböck G (duben 2005). "Dálkové ovládání chování prostřednictvím geneticky cílené fotostimulace neuronů". Buňka. 121 (1): 141–52. doi:10.1016 / j.cell.2005.02.004. PMID 15820685.
- ^ Zhang F, Wang LP, Boyden ES, Deisseroth K (říjen 2006). "Channelrhodopsin-2 a optická kontrola excitabilních buněk". Nat. Metody. 3 (10): 785–92. doi:10.1038 / nmeth936. PMID 16990810.
- ^ Lin JY, Lin MZ, Steinbach P, Tsien RY (březen 2009). "Charakterizace vytvořených variant kanálrhodopsinu s dalšími vylepšenými fotoproudy a kinetikou". Biophys. J. 96 (5): 1803–14. doi:10.1016 / j.bpj.2008.11.034. PMC 2717302. PMID 19254539.
- ^ Adamantidis AR, Zhang F, Aravanis AM, Deisseroth K, de Lecea L (listopad 2007). "Neurální substráty probuzení sondované s optogenetickou kontrolou hypocretinových neuronů". Příroda. 450 (7168): 420–4. doi:10.1038 / nature06310. PMC 6744371. PMID 17943086.
- ^ Tsai HC, Zhang F, Adamantidis A, Stuber GD, Bonci A, de Lecea L, Deisseroth K (květen 2009). „Fázová palba v dopaminergních neuronech je dostatečná pro kondicionování chování“. Věda. 324 (5930): 1080–4. doi:10.1126 / science.1168878. PMC 5262197. PMID 19389999.
- ^ Gradinaru V, Mogri M, Thompson KR, Henderson JM, Deisseroth K (duben 2009). „Optická dekonstrukce parkinsonských nervových obvodů“. Věda. 324 (5925): 354–9. CiteSeerX 10.1.1.368.668. doi:10.1126 / science.1167093. PMC 6744370. PMID 19299587.
- ^ Kravitz AV, Freeze BS, Parker PR, Kay K, Thwin MT, Deisseroth K, Kreitzer AC (červenec 2010). „Regulace parkinsonského motorického chování pomocí optogenetické kontroly obvodů bazálních ganglií“. Příroda. 466 (7306): 622–6. doi:10.1038 / nature09159. PMC 3552484. PMID 20613723.
- ^ Lee JH, Durand R, Gradinaru V, Zhang F, Goshen I, Kim DS, Fenno LE, Ramakrishnan C, Deisseroth K (červen 2010). „Globální a místní signály fMRI řízené neurony definovanými optogeneticky podle typu a zapojení“. Příroda. 465 (7299): 788–92. doi:10.1038 / nature09108. PMC 3177305. PMID 20473285.
- ^ A b Kätzel D, Zemelman BV, Buetfering C, Wölfel M, Miesenböck G (leden 2011). „Sloupcovitá a laminární organizace inhibičních spojení s neokortikálními excitačními buňkami“. Nat. Neurosci. 14 (1): 100–7. doi:10.1038 / č. 2687. PMC 3011044. PMID 21076426.
- ^ Wang H, Peca J, Matsuzaki M, Matsuzaki K, Noguchi J, Qiu L, Wang D, Zhang F, Boyden E, Deisseroth K, Kasai H, Hall WC, Feng G, Augustine GJ (květen 2007). „Vysokorychlostní mapování synaptické konektivity pomocí fotostimulace u transgenních myší Channelrhodopsin-2“. Proc. Natl. Acad. Sci. USA. 104 (19): 8143–8. doi:10.1073 / pnas.0700384104. PMC 1876585. PMID 17483470.
- ^ Mohanty SK, Reinscheid RK, Liu X, Okamura N, Krasieva TB, Berns MW (říjen 2008). „Hloubková aktivace excitabilních buněk senzibilizovaných na kanál 2-rhodopsin s vysokým prostorovým rozlišením pomocí dvoufotonové excitace laserem s blízkým infračerveným paprskem“. Biophys. J. 95 (8): 3916–26. doi:10.1529 / biophysj.108.130187. PMC 2553121. PMID 18621808.
- ^ Rickgauer JP, Tank DW (září 2009). „Dvoufotonová excitace kanálu rhodopsinu-2 při nasycení“. Proc. Natl. Acad. Sci. USA. 106 (35): 15025–30. doi:10.1073 / pnas.0907084106. PMC 2736443. PMID 19706471.
- ^ Andrasfalvy BK, Zemelman BV, Tang J, Vaziri A (červen 2010). „Dvoufotonová jednobuněčná optogenetická kontrola neuronální aktivity tvarovaným světlem“. Proc. Natl. Acad. Sci. USA. 107 (26): 11981–6. doi:10.1073 / pnas.1006620107. PMC 2900666. PMID 20543137.
- ^ Reiner A, Isacoff EY (říjen 2013). „Cena Brain 2013: optogenetická revoluce“. Trendy v neurovědách. 36 (10): 557–60. doi:10.1016 / j.tins.2013.08.005. PMID 24054067.
Další čtení
- Hegemann P. (2008). "Senzorické fotoreceptory řas". Annu Rev Plant Biol. 59: 167–189. doi:10.1146 / annurev.arplant. 59.032607.092847. PMID 18444900. (Naturel funkce channelrhodopsins a jiných fotoreceptorů zeleně)
- Arenkiel BR, Peca J, Davison IG a kol. (Duben 2007). „In vivo světelně indukovaná aktivace nervových obvodů u transgenních myší exprimujících channelrhodopsin-2“. Neuron. 54 (2): 205–18. doi:10.1016 / j.neuron.2007.03.005. PMC 3634585. PMID 17442243. (Použití channelrhodopsinu u transgenních myší ke studiu mozkových obvodů)
- Bi A, Cui J, Ma YP a kol. (Duben 2006). „Ektopická exprese rhodopsinu mikrobiálního typu obnovuje vizuální reakce u myší s degenerací fotoreceptorů“. Neuron. 50 (1): 23–33. doi:10.1016 / j.neuron.2006.02.026. PMC 1459045. PMID 16600853. (Potenciální použití channelrhodopsinu k léčbě slepoty)
externí odkazy
- OpenOptogenetics.org, komplexní wiki o optogenetice.
- Centrum optogenetiky / laboratoř Deisseroth
- Boyden laboratoř
- Laboratoř Zhuo-Hua Pan
- Laboratoř Hegemann
- Cena za mozek 2013 pro vynález optogenetiky