Bothriolepis - Bothriolepis
| Bothriolepis | |
|---|---|
 | |
| Model B. canadensis | |
| Vědecká klasifikace | |
| Království: | Animalia |
| Kmen: | Chordata |
| Třída: | †Placodermi |
| Objednat: | †Antiarchi |
| Rodina: | †Bothriolepididae |
| Rod: | †Bothriolepis Eichwald, 1840 |
| Druh | |
| |
Bothriolepis (řecký: „pitted scale“ nebo „trench scale“) byl rozšířený, hojný a různorodý rod z antiarch placoderms který žil během Střední na Pozdní devon období Paleozoická éra. Historicky, Bothriolepis sídlil v řadě paleo prostředí rozložených na každém paleokontinentu, včetně pobřežních mořských a sladkovodních prostředí.[1] Většina druhů Bothriolepis byly charakterizovány jako relativně malé, bentický, sladkovodní detritivores (organismy, které získávají živiny konzumací rozkládajícího se rostlinného / živočišného materiálu), v průměru kolem 30 centimetrů (12 palců) na délku.[2] Největší druh, B. rex, měl odhadovanou délku těla 170 centimetrů (67 palců). Ačkoli je rozsáhlý s více než 60 druhy nalezenými po celém světě,[3] poměrně Bothriolepis není neobvykle rozmanitější než většina moderních druhů žijících na dně v dnešní době.[4]
Klasifikace
Bothriolepis je rod umístěný v řádu placodermů Antiarchi. Nejstarší antiarchové plakodermy se poprvé objevily v Silurian období paleozoické éry a lze je nalézt distribuované na každém paleokontinent devonským obdobím.[5] Nejčasnější členové Bothriolepis objeví se středním devonem. Antiarchové, stejně jako další placodermové, jsou morfologicky různorodí a vyznačují se kostnatými deskami, které zakrývají jejich hlavu a přední část kmene.[5] Raná ontogenetická stadia placodermů měla tenčí kostní destičky jak v hlavě, tak i v kufru, což umožňovalo snadné rozlišení mezi časnými ontogenetickými stadii placoderma ve fosilním záznamu a taxony, které vlastnily plně vyvinuté kostní destičky, ale byly podle charakterizace malé.[5] Kostní desky Placoderm byly obecně tvořeny třemi vrstvami, včetně kompaktní bazální lamelární kostní vrstvy, střední houbovité kostní vrstvy a povrchové vrstvy;[5] Bothriolepis může být klasifikován jako placoderm, protože má tyto vrstvy. Na konci devonu placodermové vyhynuli.[5] Placodermi je paraphyletic skupina kladu Gnathostomata, který zahrnuje všechny čelistní obratlovce.[5] Není jasné, kdy přesně se gnathostomy objevily, ale skromný časný fosilní záznam naznačuje, že to bylo někdy v rané paleozoické éře.[6] Poslední druh Bothriolepis zemřel spolu se zbytkem Placodermi na konci devonského období.
Obecná anatomie

Hlava
V hlavě jsou dva otvory Bothriolepis: otvor pro klíčovou dírku podél středové čáry na horní straně pro oči a nosní dírky a otvor pro ústa na spodní straně blízko předního konce hlavy. Objev týkající se zachovaných struktur, které se zdají být nosními kapslemi, potvrzuje přesvědčení, že vnější nosní otvory leží na hřbetní straně hlavy blízko očí.[7] Navíc je poloha úst na ventrální straně lebky v souladu s typickou horizontální klidovou orientací Bothriolepis. To mělo zvláštní rys na jeho lebce, samostatnou přepážku kosti pod otvorem pro oči a nosní dírky obklopující nosní tobolky, nazývané preorbitální výklenek.
Čelist
Nový vzorek z Gogo formace v Canning Basin v západní Austrálii poskytl důkazy týkající se morfologických rysů viscerálních čelistních prvků Bothriolepis. Pomocí vzorku je zřejmé, že mentální ploténka (dermální kost, která tvoří horní část čelisti) antiarchů je homologní se suborbitální ploténkou nalezenou u jiných placodermů. Dolní čelist se skládá z diferencovaných čepelí a kousacích částí. Vedle mandibulárního kloubu jsou prelaterální a infraprelaterální dlahy, které jsou obě kosti nesoucí kanál. The palatoquadrate postrádá vysoký orbitální proces a byl připevněn pouze k ventrální části mentální ploténky, což dokazuje, že etmoidální oblast mozkové skříně (oblast lebky oddělující mozek a nosní dutinu) byla ve skutečnosti hlubší, než se původně věřilo.[8] Kromě výše uvedeného vzorku z formace Gogo bylo nalezeno několik dalších vzorků s ústními částmi drženými v přirozené poloze membránou, která pokrývá ústní oblast a připojuje se k bočním a předním okrajům hlavy.[9] Bothriolepis má čelist, ve které jsou obě poloviny oddělené a u dospělých jsou funkčně nezávislé.[9]
Kmen

Bothriolepis měl štíhlý kufr, který byl pravděpodobně pokryt měkkou kůží bez šupin nebo značek. Orientace, která se zdá být nejstabilnější pro odpočinek, byla hřbetní plocha nahoru, o čemž svědčí plochý povrch na ventrální straně.[1] Obrys kufru naznačuje, že tam mohla být a notochord přítomný obklopený membránovým obalem,[9] o tom však neexistují žádné přímé důkazy, protože notochord je tvořen měkkou tkání, která není ve fosilním záznamu typicky zachována. Podobně jako ostatní antiarchy, hrudní štít Bothriolepis byl připevněn k jeho těžce obrněné hlavě. Jeho krabicovité tělo bylo uzavřeno v pancéřových deskách, které poskytovaly ochranu před predátory. K ventrálnímu povrchu kmene je připevněna velká, tenká, kruhová deska vyznačená hluboko položenými liniemi a povrchovými hřebeny. Tato deska leží těsně pod otvorem do kloaka.[9]
Dermální kostra
Dermální kostra je organizována ve třech vrstvách: povrchová lamelová vrstva, spongiosa spongiosa a kompaktní bazální lamelová vrstva. Dokonce i na počátku ontogeneze jsou tyto vrstvy patrné ve vzorku Bothriolepis canadensis. Nejprve se vyvinou kompaktní vrstvy.[10] Předpokládá se, že povrchová vrstva má denticly, které mohou být vyrobeny z buněčné kosti.[11]
Ploutve a ocas

Bothriolepis měl dlouhý pár podobný páteři prsní ploutve, připojil se k základně a znovu o něco více než v polovině. Tyto špice podobné ploutve byly pravděpodobně použity ke zvednutí těla ze dna; díky jeho těžkému brnění by se rychle potopilo, jakmile ztratilo hybnost.[2][12] Může také použít své prsní ploutve k tomu, aby přes sebe házel sediment (bláto, písek nebo jiné). Kromě prsních ploutví měla také dvě hřbetní ploutve: nízkou, podlouhlou přední hřbetní ploutev a vysokou zaoblenou zadní hřbetní ploutev [9]- ačkoli předpokládaná struktura hřbetních ploutví se liší podle konkrétního druhu Bothriolepis a byl několikrát upraven v rekonstrukcích uvolněných vědci, jakmile byly k dispozici nové informace. The ocasní ocas byl protáhlý, končící úzkým pásmem, ale bohužel je zřídka zachován ve fosiliích.[9] Ačkoli neexistuje dohodnuté vysvětlení jejich funkce, Bothriolepis měl také dva membránové ventrální límce umístěné na zadním konci trupu krunýře na obou stranách ocasu, z nichž každý měl dvě odlišné oblasti.[7] Neexistují žádné důkazy o tom, že límce se podílely na podpoře kostry, ale je možné, že buď fungovaly jako ploutve, nebo se podílely na reprodukci, a mohly být dokonce přítomny u jednoho pohlaví, u druhého nikoli.[7]
Měkká anatomie

Struktury složené z měkké tkáně se ve fosiliích obvykle nezachovávají, protože se snadno rozkládají a rozkládají mnohem rychleji než tvrdé tkáně, což znamená, že ve fosilním záznamu často chybí informace týkající se vnitřní anatomie fosilních druhů. K ochraně struktur měkkých tkání může někdy dojít, pokud sedimenty vyplní vnitřní struktury organismu po jeho smrti nebo po ní. Papír Roberta Denisona s názvem „The Soft Anatomy of Bothriolepis"zkoumá formy a orgány Bothriolepis.[7] Tyto vnitřní struktury byly zachovány, když různé typy sedimentů obklopujících exteriér zvířete vyplňovaly vnitřní krunýře (tímto způsobem bylo možné zachovat pouze orgány, které komunikují s exteriérem). V různých částech systému byly identifikovány tři různé typy sedimentů Bothriolepis: první bledě zelenošedý středně strukturovaný pískovec převážně sestávající z kalcitu; druhý podobný, ale jemnější sediment, který zachovává mnoho forem orgánů; a třetí zřetelný jemnozrnný prachovec skládající se z křemene, slídy a dalších minerálů, ale bez kalcitu.[7] Tyto sedimenty pomohly zachovat následující vnitřní prvky:
Zažívací systém
Obecně platí, že zažívací systém z Bothriolepis –Který zahrnuje orgány podílející se na požití, trávení a odstraňování odpadu - lze popsat jako jednoduché a přímé, na rozdíl od lidských. Začíná to na předním konci organismu s malou ústní dutinou umístěnou nad zadní oblastí desek horní čelisti. Zezadu od úst se zažívací systém rozšiřuje do širší a dorzoventrálně zploštělé oblasti zvané hltan, ze kterých vycházejí žábry i plíce. The jícen, který je také charakterizován jako dorso-ventrálně zploštělá trubice, sahá od úst do žaludku a vede k zploštělé elipsoidní struktuře. Tato struktura může být homologní s předním koncem střeva nalezeným u jiných ryb.[7] Plochost těchto struktur mohla být přehnaná, když fosilní vzorky zažily tektonická deformace v geologickém čase. Střevo začíná úzce na předním konci, rozpíná se příčně a pak se znovu zužuje dozadu směrem k válcovitému konečníku, který končí těsně za zadním koncem krunýře trupu. I když je zažívací systém primitivní povahy a postrádá rozšířenou oblast žaludku, specializuje se na samostatně získávanou komplexní spirálovou chlopní, srovnatelnou s elasmobranchs a mnoho kostnatých ryb a podobné těm, které se vyskytují u některých žraloků. Jediný záhyb tkáně navinutý na vlastní osu tvoří tento specializovaný spirálový ventil.[7]
Žábry
Odvozuje se, že žábry Bothriolepis jsou primitivního typu, i když jejich struktura stále není dobře pochopena. Postranně jsou uzavřeny operulárním záhybem a nacházejí se v prostoru pod boční částí štítu hlavy, rozprostírající se mediálně pod neurocranium. Ve srovnání s žábry normálně tvarovaných ryb je oblast žábry z Bothriolepis je považován za umístěný více dorzálně, je vpředu více přeplněný a obecně je relativně krátký a široký.[7]
Spárované ventrální vaky


Zadní část z krunýře trupu se táhnou spárované ventrální vaky, které sahají až k přednímu konci spirálového střeva. Zdá se, že vaky pocházejí z hltanu jako jedna střední trubice, která se poté rozšiřuje dozadu a nakonec se rozdělí na dva vaky, které mohou být homologní s plíce jisté dipnoans a tetrapody.[7] Předpokládalo se, že tyto plíce, spojené s kloubovými pažemi a pevnou podpůrnou kostrou, by umožnily Bothriolepis cestovat na souši. Navíc jako Robert Denison[7] uvádí, protože neexistují žádné důkazy o spojení mezi externím naris a ústa, Bothriolepis pravděpodobně dýchal podobně jako dnešní lungfish, tj. umístěním úst nad vodní hladinu a spolknutím vzduchu.
Navzdory původní interpretaci, kterou předložil Denison v roce 1941, ne všichni paleontologové souhlasí, že placodermům se líbí Bothriolepis ve skutečnosti vlastnil plíce. Například v jeho příspěvku „Plíce“ u placodermů, perzistentní paleobiologický mýtus související s interpretacemi předsudků o životním prostředíD. Goujet naznačuje, že i když ze sedimentárních struktur mohou být patrné stopy některých zažívacích orgánů, neexistují žádné důkazy podporující přítomnost plic ve vzorcích z kanadské formace Escuminac, na nichž bylo založeno původní tvrzení. Poznamenává, že celosvětová distribuce Bothriolepis je omezena na přísně mořském prostředí, a proto věří, že přítomnost plic v Bothriolepis je nejistý. Pravděpodobně je nutné další vyšetřování fosilií, aby se dospělo k závěru o přítomnosti plic v Bothriolepis.[13]
Krmení
Bothriolepis, stejně jako u všech ostatních antiarchů, se předpokládá, že se krmili přímým polykáním plných bláta a jiných měkkých sedimentů, aby se strávili detritus, malé mikroorganismy, řasy a jiné formy organických látek v polykaných sedimentech. Navíc to dále naznačuje umístění úst na ventrální straně hlavy Bothriolepis byl pravděpodobně spodní krmítko. Pravidelná přítomnost „uhlíkatého materiálu v zažívacím traktu“ naznačuje, že většinu jeho potravy tvořil rostlinný materiál.[7]
Rozdělení
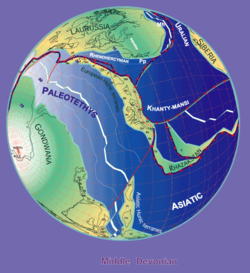
Autor: Stampfli & Borel, 2000
Bothriolepis fosilie se nacházejí ve střední a pozdní Devonský vrstvy (před 387 až 360 miliony let).[12] Protože fosilie se nacházejí ve sladkovodních sedimentech, Bothriolepis se předpokládá, že strávil většinu svého života v sladkovodní řek a jezer, ale pravděpodobně byl schopen vstoupit slaná voda také proto, že se zdálo, že jeho rozsah odpovídal devonským kontinentálním pobřežím. Velká seskupení Bothriolepis vzorky byly nalezeny v Asii, Evropě, Austrálii (Gogo formace ),[9] Pensylvánie (formace Catskill),[1] Quebec (Formace Escuminac ),[1] Virginie (Chemung),[14] Colorado,[14] Cuche formace (Boyacá, Kolumbie),[15] a po celém světě.
Místo vytvoření Catskill
The Catskill formace (Svrchní devon, Famennian Stage), umístěný v Okres Tioga, Pensylvánie, je místem velkého vzorku malých jedinců Bothriolepis. Vzorek byl odebrán ze série kamenných desek, které se skládaly z částečných nebo úplných, kloubových, vnějších skeletů. Bylo nalezeno více než dvě stě jedinců zabalených těsně vedle sebe s malým až žádným překrytím. Z tohoto vzorku mnoho informací o charakteristikách mladistvých Bothriolepis lze určit. Morfometrická studie provedená Jasonem Downsem a spoluautoři zdůrazňuje určité vlastnosti, které naznačují mladistvost Bothriolepis, včetně středně velké hlavy a středně velké orbitální fenestra — Obojí jsou charakteristiky, které nejmenší uznal také Erik Stensio v roce 1948 B. canadensis Jednotlivci.[1] Ve vzorku Catskill je také vidět několik dalších rysů, které Stensio označil jako indikativní pro mladé jedince. Mezi tyto vlastnosti patří „jemné dermální kosti s ornamentem skládající se spíše z kontinuálních anastomózních hřebenů než tuberkul, hřbetní štít kmene užší než dlouhý a s nepřetržitým a výrazným hřbetním středním hřebenem a předmediánní dlahou, která je širší než dlouhá“.[1]
Druh
Paleontologie obratlovců je silně závislá na schopnosti rozlišovat mezi různými druhy způsobem, který je konzistentní jak v rámci konkrétního rodu, tak ve všech organismech. Rod Bothriolepis není výjimkou z této zásady. Níže je uvedeno několik významných druhů uvnitř Bothriolepis; celkem bylo pojmenováno více než šedesát druhů a je pravděpodobné, že značná část z nich je platná kvůli kosmopolitní povaze Bothriolepis.[3]
Bothriolepis canadensis

Bothriolepis canadensis je taxon, který často slouží jako modelový organismus řádu Antiarchi kvůli svému obrovskému vzorku úplných, neporušených vzorků nalezených na Formace Escuminac v Quebec, Kanada.[1] Vzhledem k obrovské velikosti vzorku se tento druh často používá k porovnání údajů o růstu nově získaných vzorků Bothriolepis, včetně těch, které se nacházejí ve výše zmíněné formaci Catskill. Toto srovnání umožňuje vědcům určit, zda nově nalezené vzorky představují mladistvé jedince nebo nové druhy „Bothriolepis“.
B. canadensis byl poprvé popsán v roce 1880 J. F. Whiteavesem s použitím omezeného počtu znetvořených vzorků. Dalším, kdo navrhl rekonstrukci tohoto druhu, byl W. Patten, který své nálezy publikoval v roce 1904 po objevu několika exemplářů, které byly dobře zachovány ve 3D. V roce 1948 vydal E. Stensio podrobné zobrazení B. canadensis anatomie s využitím množství materiálu, který se nakonec stal nejuznávanějším popisem tohoto druhu. Od publikace Stensio mnoho dalších poskytlo rekonstruované modely B. canadensis s upravenými aspekty anatomie, včetně Vezinovy modifikované jediné hřbetní ploutve a nověji, rekonstrukce od Arsenault et al ze vzorků s malým taphonomickým zkreslením. V současné době model Arsenault et al. je považován za nejpřesnější, zatímco stále existuje mnoho debat o různých aspektech vnější anatomie tohoto druhu. Přes nejistotu B. canadensis je stále klasicky považován za jeden z nejznámějších druhů.[16]
Externí kostra z Bothriolepis canadensis je vyroben z buněčné dermální kostní tkáně a vyznačuje se odlišnou horizontální zonací nebo stratifikací.[10] Modelka má průměrnou celkovou délku 43,67 centimetrů (17,19 palce) a průměrnou délku dermálního pancíře 15,53 centimetru (6,11 palce), což představuje 35,6% odhadované celkové délky.[16] Jako mnoho antiarchů, B. canadensis měl také úzké prsní ploutve, a heterocercal ocasní ploutev (což znamená, že notochord zasahuje do horního laloku ocasní ocasu) a velká hřbetní ploutev, která pravděpodobně nehrála důležitou roli v pohonu, ale místo toho působila spíše jako stabilizátor [16]
Bothriolepis rex
Původně popsali Downs et al. (2016), Bothriolepis rex pochází z formace Nordstrand Point na ostrově Ellesmere v Kanadě. Délka těla B. rexe se odhaduje na 1,7 metru, a je tedy největším známým druhem Bothriolepis. Jeho brnění je obzvláště silné a husté, i když vezmeme v úvahu jeho velikost. Downs a kol. (2016) naznačují, že to mohlo chránit zvíře před velkými predátory a sloužit jako zátěž, aby se zabránilo tomuto velkému obyvateli dna vznášet se na povrch.
Bothriolepis virginiensis


Původně popsali Weems et al. v roce 1981 tento druh, Bothriolepis virginiensis, pochází z „Chemung“ poblíž Winchester, Virginie. Několik znaků nalezených v B. virginiensis lze nalézt také u jiných druhů Bothriolepis, (zvláště B. nitida), včetně zadních šikmých hlavových drážek smyslové linie, které se setkávají relativně daleko vpředu na šíjové dlaze, relativně protáhlého orbitálního fenestru a nízkého předního-středního-hřbetního hřebenu.[14] Charakteristiky, které rozlišují B. virginiensis z jiných druhů zahrnují, ale nejsou na ně omezeny, fúzované stehy hlavy, fúzované prvky v segmentech distální prsní ploutve dospělých a dlouhá premediánová destička vzhledem k délce čelního štítu.[14]
V současné době existuje mnoho debat o tom, zda druh B. virginiensis a B. nitida lze ve skutečnosti od sebe odlišit. Thomson a Thomas uvádějí, že pět druhů Bothriolepis ze Spojených států (B. nitida, B. minor, B. virginiensis, B. darbiensis a B. coloradensis) nelze navzájem důsledně odlišit.[4] Weems naopak tvrdí, že existuje několik zvláštností, které odlišují druhy od sebe, včetně několika výše uvedených.[14]
Bothriolepis nitida
Tento druh, nalezený v dnešní době Pensylvánie, byl původně popsán J. Leidym v roce 1856. Jak bylo uvedeno výše, existuje mnoho debat o rozlišitelnosti mezi B. nitida a B. virginiensis, avšak na základě důkazů předložených Weemsem (2004),[14] pro každý druh existuje několik odlišitelných zvláštností. B. nitida má maximální délku čelního štítu 65 milimetrů (2,6 palce), úzký a mělký trifidní preorbitální výklenek, přední a střední dorzální (AMD) dlahu, která je širší než dlouhá, a ventrální hrudní štít s konvexními bočními okraji.[14]
Bothriolepis coloradensis
Poprvé popsán Eastmanem v roce 1904, tento druh byl nalezen lokalizovaný v dnešní době Colorado. Existuje možnost, že tento druh je podobný, ne-li totožný B. nitida, nicméně proto, že materiál je k dispozici B. coloradensis je roztříštěný, je nemožné oba druhy s jistotou srovnávat.[14]
Bothriolepis africana
Bothriolepis africana [17] je Bothriolepis druhy známé z nejvyšších paleolatitude, popsáno z vkladů původně uložených v rámci Pozdní devon Antarktický kruh. Zbytky byly získány výhradně z jedné uhlíkaté břidlice poblíž vrcholu pozdního devonu, Famennian, Formace Witpoort (skupina Witteberg) vystavená na silnici vedoucí jižně od Makhandy /Grahamstown v Jižní Africe. Tato stránka je Waterloo Farm lagerstätte je interpretován jako představující zadní bariéru pobřežního lagunálního prostředí s mořskými i fluviálními vlivy.[18] Gess to pozoroval Bothriolepis byl v lokalitě Waterloo Farm méně hojný než nanejvýš Bothriolepis- ložiskové lokality, ačkoli je zastoupena celá ontogenetická řada. Délka pancíře hlavy a trupu se pohybovala mezi 20 a 300 mm, což se promítá na základě proporcí dvou nejmenších jedinců (u nichž jsou zachovány otisky ocasu) do délek celého těla pohybujících se mezi 52 a 780 mm.[19]Podle [20] Bothriolepis africana byl považován za nejvíce podobný Bothriolepis barretti [21] od pozdních Givetian z Antarktida. Podobnosti mezi nimi byly použity k navržení odvození Bothriolepis africana z východu Gondwanan životní prostředí. [22]


Reference
- ^ A b C d E F G Downs, J.P .; Criswell, K.E .; Daeschler, E.B. (Říjen 2011). „Masová úmrtnost mladistvých antiarchů (Bothriolepis sp.) z formace Catskill (svrchní devon, fáze Famennian), okres Tioga, Pensylvánie “. Sborník Akademie přírodních věd ve Filadelfii (161): 191–203.
- ^ A b Palmer, D., ed. (1999). Marshallova ilustrovaná encyklopedie dinosaurů a prehistorických zvířat. London: Marshall Editions. str. 33. ISBN 978-1-84028-152-1.
- ^ A b Young, G.C. (2010). „Placoderms (Armored Fish): Dominant Vertebrates of the Devonian Period“. Výroční přehled o Zemi a planetárních vědách. 38: 523–550. doi:10.1146 / annurev-earth-040809-152507.
- ^ A b Thomson, K. S.; Thomas, B. (srpen 2001). „O stavu druhů Bothriolepis (Placodermi, Antiarchi) v Severní Americe “. Časopis paleontologie obratlovců. 21 (4): 679–686. doi:10.1671 / 0272-4634 (2001) 021 [0679: otsoso] 2.0.co; 2.
- ^ A b C d E F Johanson, Zerina; Trinajstic, Kate (2014). „Fosilizované ontogeneze: příspěvek ontogeneze placodermu k našemu chápání vývoje raných gnathostomů“. Paleontologie. 57 (3): 505–516. doi:10.1111 / pala.12093.
- ^ Brazeau, M. (2009). „Mozková skříň a čelisti devonského„ akantodiana “a moderní původ gnathostome“ (PDF). Příroda. 457 (7227): 305–308. doi:10.1038 / nature07436. hdl:10044/1/17971. PMID 19148098.
- ^ A b C d E F G h i j k Denison, R.H. (září 1941). "Měkká anatomie Bothriolepis". Journal of Paleontology. 15 (5): 553–561.
- ^ Young, G.C. (1984). „Rekonstrukce čelistí a mozkovny v devonských placodermových rybách Bothriolepis". Paleontologie. 27 (3): 635–661.
- ^ A b C d E F G Patten, W. (červenec 1904). "Nová fakta týkající se Bothriolepis". Biologický bulletin. 7 (2): 113–124. doi:10.2307/1535537. JSTOR 1535537.
- ^ A b Downs, J.P .; Donoghue, P.C.J. (2009). "Skeletální histologie Bothriolepis canadensis (Placodermi, Antiarchi) a vývoj kostry u původu čelistních obratlovců “. Journal of Morphology. 270 (11): 1364–1380. doi:10,1002 / jmor.10765. PMID 19533688.
- ^ Giles, S. (2013). „Histologie dermálních koster„ placoderm “: důsledky pro povahu rodového gnathostomu“. Journal of Morphology. 274 (6): 627–644. doi:10.1002 / jmor.20119. PMC 5176033. PMID 23378262.
- ^ A b "Muzeum stáří ryb - fosilie". Muzeum Age of Fishes, Nový Jižní Wales, Austrálie.
- ^ Goujet, D. (2011). ""Plíce „u placodermů, přetrvávající paleobiologický mýtus související s předpojatými interpretacemi prostředí“. Comptes Rendus Palevol. 10 (5–6): 323–329. doi:10.1016 / j.crpv.2011.03.008.
- ^ A b C d E F G h Weems, R.E. (Březen 2004). "Bothriolepis viginiensis, platný druh placodermových ryb oddělitelných od Bothriolepis nitida". Časopis paleontologie obratlovců. 24 (1): 245–250. doi:10.1671/20.
- ^ Janvier, Philippe; Villarroel A, Carlos (1998). „Los Peces Devónicos del Macizo de Floresta (Boyacá, Kolumbie). Consideraciones taxonómicas, bioestratigráficas, biogeográficas y ambientales“. Geología Colombiana. 23: 3–18. Citováno 2017-03-31.
- ^ A b C Bechard, I .; Arsenault, F .; Cloutier, R .; Kerr, J. (2014). „Devonská ryba Bothriolepis canadensis znovu s trojrozměrnými digitálními snímky “. Palaeontologia Electronica. 17 (1).
- ^ LONG, J. A., ANDERSON, M. E., GESS, R. W. & HILLER, N. (1997). Nové placodermové ryby z pozdního devonu v Jižní Africe. Časopis paleontologie obratlovců 17 253–268.
- ^ Gess, Robert W .; Whitfield, Alan K. (14. února 2020). „Vývoj ústí ryb a tetrapodů: poznatky z pozdně devonského (famennského) gondwanského jezera v ústí řek a jihoafrického ekvivalentu holocénu“. Biologické recenze. doi: 10,1111 / brv.12590. PMID 32059074
- ^ GESS, R.W. (2011). Gondwananské fameniánské vzorce biodiverzity s vysokou šířkou - důkazy z jihoafrického souvrství Witpoort (Cape Supergroup, WittebergGroup). Diplomová práce: University of the Witwatersrand, Johanneburg.
- ^ LONG, J. A., ANDERSON, M. E., GESS, R. W. & HILLER, N. (1997). Nové placodermové ryby z pozdního devonu v Jižní Africe. Časopis paleontologie obratlovců 17 253–268.
- ^ YOUNG, G. (1984). Rekonstrukce čelistí a mozkovny v devonském placodermu postihuje Bothriolepis. Paleontologie 27, 635–661.
- ^ LONG, J. A., ANDERSON, M. E., GESS, R. W. & HILLER, N. (1997). Nové placodermové ryby z pozdního devonu v Jižní Africe. Časopis paleontologie obratlovců 17 253–268.
externí odkazy
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||

