Trofická soudržnost - Trophic coherence
Trofická soudržnost je majetkem řízené grafy (nebo režie sítí ).[1] Je založen na konceptu trofické úrovně používá se hlavně v ekologie,[2] ale které lze obecně definovat pro směrované sítě a poskytuje míru hierarchické struktury mezi uzly. Trofická koherence je tendence uzlů spadat do přesně definovaných trofických úrovní. Souvisí to s několika strukturálními a dynamickými vlastnostmi směrovaných sítí, včetně prevalence cykly[3] a síťové motivy,[4] ekologická stabilita,[1] interaktivita,[5] a šíření procesů jako epidemie a neuronové laviny.[6]
Definice
Zvažte směrovanou síť definovanou matice sousedství .Každý uzel lze přiřadit a tropická úroveň podle





kde je je ve stupních a uzly s (bazální uzliny) mají podle konvence. Každá hrana má a trofický rozdíl přidružené, definované jako .v trofická soudržnost sítě je měřítkem toho, jak pevně vyvrcholila distribuce trofických vzdáleností,, je kolem své střední hodnoty, která je vždy To může zachytit parametr nesoudržnosti , rovnající se směrodatné odchylce :








kde je počet hran v síti.[1]

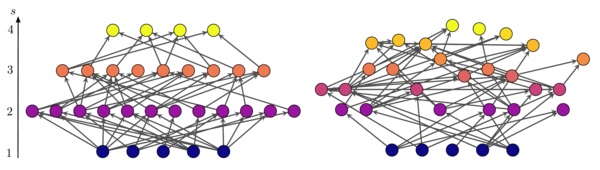
Obrázek ukazuje dvě sítě, které se liší svou trofickou koherencí. Poloha uzlů na svislé ose odpovídá jejich trofické úrovni. V síti nalevo spadají uzly do odlišných (celých) trofických úrovní, takže síť je maximálně koherentní . V jednom napravo má mnoho uzlů dílčí trofické úrovně a síť je více nesouvislá .[6]


Trofická soudržnost v přírodě
Míru, do jaké jsou empirické sítě troficky koherentní (nebo nekoherentní), lze zjistit srovnáním s nulovým modelem. bazální soubor, který zahrnuje sítě, ve kterých mají všechny ne-bazální uzly stejný podíl bazálních uzlů u sousedů.[3] Očekávané hodnoty v tomto souboru se sbližují s těmi široce používanými konfigurační soubor[7] v limitu , (s a počty uzlů a hran) a lze je numericky zobrazit jako dobrou aproximaci pro konečné náhodné sítě. Očekávání bazálního souboru pro parametr nesoudržnosti je





kde je počet hran připojených k bazálním uzlům.[3]Poměr měřeno v empirických sítích odhaluje, zda jsou víceméně koherentní než náhodné očekávání. Například Johnson a Jones[3] najít v sadě sítí, které potravinářské weby jsou výrazně koherentní , metabolické sítě jsou výrazně nesouvislé , a genové regulační sítě jsou blízké náhodnému očekávání .





Trofické úrovně a funkce uzlu
Dosud není dostatečné pochopení mechanismů, které by mohly vést k tomu, že se určité druhy sítí stanou výrazně koherentními nebo nesoudržnými.[3] V systémech, které představují korelace mezi trofickou úrovní a dalšími vlastnostmi uzlů, by však procesy, které měly tendenci upřednostňovat vytváření hran mezi uzly se zvláštními charakteristikami, mohly vyvolat soudržnost nebo nesoudržnost. V případě potravinových sítí se predátoři obvykle specializují na konzumaci kořisti s určitými biologickými vlastnostmi (jako je velikost, rychlost nebo chování), které korelují s jejich stravou, a tedy s trofickou úrovní. To bylo navrženo jako důvod pro soudržnost potravinového webu.[1] Modely potravinářského webu založené na a osa výklenku nereprodukujte realistickou trofickou soudržnost,[1] což může znamenat, že toto vysvětlení není dostatečné, nebo že jich je několik rozměry výklenku je třeba zvážit.[8]

Vztah mezi trofickou úrovní a funkcí uzlu lze vidět v jiných sítích než v potravinářských sítích. Obrázek ukazuje síť slovních sousedů odvozenou z knihy Zelená vejce a šunka tím, že Dr. Seuss.[3] Výška uzlů představuje jejich trofické úrovně (zde podle směru okraje, který je opakem směru naznačeného šipkami, které označují pořadí, ve kterém jsou slova ve větách zřetězena). Syntaktická funkce slov je také zobrazena barvou uzlu. Existuje jasný vztah mezi syntaktickou funkcí a trofickou úrovní: střední trofická úroveň běžných podstatných jmen (modrá) je , zatímco u sloves (červená) je Tento příklad ukazuje, jak může z funkce uzlu vyplynout tropická soudržnost nebo nesoudržnost, a také to, že trofická struktura sítí poskytuje prostředek k identifikaci funkce uzlu v určitých systémech.


Generování troficky koherentních sítí
Existují různé způsoby generování směrovaných sítí se specifickou trofickou koherencí, vše založené na postupném zavádění nových hran do systému takovým způsobem, že pravděpodobnost přijetí každé nové kandidátské hrany závisí na očekávaném trofickém rozdílu, který by měl.
The preferenční model kořisti je vyvíjející se síťový model podobný Barábasi-Albertův model preferenčního připoutání, ale inspirován ekosystémem, který roste imigrací nových druhů.[1]Jeden začíná bazální uzly a pokračuje v zavádění nových uzlů až do celkové výše .Každý nový uzel je přidělen první soused (druh kořisti v kontextu potravinového webu) a nový okraj je umístěn z na . Nový uzel má dočasnou trofickou úroveň .Pak další noví sousedé jsou vybrány pro od těch v síti podle jejich trofických úrovní. Konkrétně pro nového kandidáta u sousedů , pravděpodobnost, že bude vybrán, je funkcí . Johnson et al[1] použití







kde je parametr, který vyladí trofickou koherenci: pro jsou generovány maximálně koherentní sítě a se zvyšuje monotónně s pro .Výběr je libovolný. Jednou z možností je nastavit na ,kde je počet uzlů, které jsou již v síti, když přijde a je náhodná proměnná získaná z a Distribuce beta s parametry a








( je požadovaný počet hran). Tímto způsobem zobecněný kaskádový model[9][10] je obnovena v limitu , a rozdělení stupňů je jako v výklenkový model[11] a zobecněný výklenkový model.[10]Tento algoritmus, jak je popsáno výše, generuje sítě bez cyklů (kromě vlastních cyklů, pokud je nový uzel je sama považována za svého kandidáta u sousedů ). Aby cykly všech délek byly možné, lze uvažovat o nových kandidátských hranách, ve kterých je nový uzel je sousedem i těmi, ve kterých by byl sousedem. Pravděpodobnost přijetí těchto hran, , pak záleží na .




The zobecněný preferenční model lovu[6] je podobný výše popsanému, ale má určité výhody. Zejména je analyticky přijatelnější a lze generovat sítě s přesným počtem hran Síť začíná bazální uzliny a pak další nové uzly jsou přidány následujícím způsobem. Když každý vstoupí do systému, je mu náhodně přiřazen jeden soused, který již existuje. Každý uzel pak má celočíselnou dočasnou trofickou úroveň . Zbývající okraje jsou zavedeny následovně. Každá dvojice uzlů má spojeny dvě dočasné trofické vzdálenosti, a . Každá z těchto kandidátních hran je přijímána s pravděpodobností, která závisí na této dočasné vzdálenosti. Klaise a Johnson[6] použití







protože shledávají rozdělení trofických vzdáleností v několika druzích sítí přibližně normální a tato volba vede k rozsahu parametru ve kterém . Jakmile byly zavedeny všechny hrany, je třeba přepočítat trofické úrovně všech uzlů, protože se budou lišit od dočasných původně přiřazených, pokud Stejně jako u preferenčního modelu lovu, parametr průměrné nesoudržnosti výsledných sítí je monotónně rostoucí funkce pro . Obrázek výše ukazuje dvě sítě s různou trofickou koherencí generované tímto algoritmem.



Reference
- ^ A b C d E F G Johnson S, Domınguez-Garcıa V, Donetti L, Muñoz MA (2014). „Trofická soudržnost určuje stabilitu potravinového webu“. Proc Natl Acad Sci USA. 111 (50): 17923–17928. arXiv:1404.7728. Bibcode:2014PNAS..11117923J. doi:10.1073 / pnas.1409077111. PMC 4273378. PMID 25468963.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Levine S (1980). "Několik opatření trofické struktury použitelné pro složité potravinové sítě". J Theor Biol. 83 (2): 195–207. doi:10.1016 / 0022-5193 (80) 90288-X.
- ^ A b C d E F Johnson S a Jones NS (2017). „Looplessness in networks is associated to trophic coherence“. Proc Natl Acad Sci USA. 114 (22): 5618–5623. arXiv:1505.07332. doi:10.1073 / pnas.1613786114. PMC 5465891. PMID 28512222.
- ^ Klaise J a Johnson S (2017). „Původ rodin motivů v potravinářských sítích“. Vědecké zprávy. 7 (1): 16197. arXiv:1609.04318. Bibcode:2017NatSR ... 716197K. doi:10.1038 / s41598-017-15496-1. PMC 5700930. PMID 29170384.
- ^ Domınguez-Garcıa V, Johnson S, Muñoz MA (2016). "Intervality a koherence ve složitých sítích". Chaos. 26 (6): 065308. arXiv:1603.03767. Bibcode:2016Chaos..26f5308D. doi:10.1063/1.4953163. PMID 27368797.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ A b C d Klaise J a Johnson S (2016). „Od neuronů po epidemie: Jak trofická koherence ovlivňuje procesy šíření“. Chaos. 26 (6): 065310. arXiv:1603.00670. Bibcode:2016Chaos..26f5310K. doi:10.1063/1.4953160. PMID 27368799.
- ^ Newman, MEJ (2003). "Struktura a funkce složitých sítí". Recenze SIAM. 45 (2): 167–256. arXiv:cond-mat / 0303516. Bibcode:2003SIAMR..45..167N. doi:10.1137 / S003614450342480.
- ^ Rossberg AG, Brännström A, Dieckmann U (2010). "Struktura potravinového webu v nízko a dimenzionálních trofických výklenkových prostorech". Rozhraní J R Soc. 7 (53): 1735–1743. doi:10.1098 / rsif.2010.0111. PMC 2988264. PMID 20462875.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Cohen JE a Newman CM (1985). „Stochastická teorie komunitních potravinových webů I. Modely a agregovaná data“. Proc. R. Soc. B. 224 (1237): 421–448. Bibcode:1985RSPSB.224..421C. doi:10.1098 / rspb.1985.0042.
- ^ A b Stouffer DB, Camacho J, Amaral LAN (2006). „Důkladná míra interakce potravinového webu“. Proc Natl Acad Sci USA. 103 (50): 19015–19020. Bibcode:2006PNAS..10319015S. doi:10.1073 / pnas.0603844103. PMC 1748169. PMID 17146055.CS1 maint: více jmen: seznam autorů (odkaz)
- ^ Williams RJ a Martinez ND (2000). "Jednoduchá pravidla přinášejí komplexní potravinové weby". Příroda. 404 (6774): 180–183. Bibcode:2000Natur.404..180W. doi:10.1038/35004572. PMID 10724169.