Terminální aminové izotopové značení substrátů - Terminal amine isotopic labeling of substrates
Terminální aminové izotopové značení substrátů (OCASY) je metoda v kvantitativní proteomika který identifikuje protein obsah vzorků na základě N-terminál fragmenty každého proteinu (N-terminální peptidy ) a detekuje rozdíly v nadbytku bílkovin mezi vzorky.
Stejně jako jiné metody založené na N-koncových peptidech používá tento test trypsin rozbít proteiny na fragmenty a oddělit N-koncové peptidy (fragmenty obsahující N-konce původních proteinů) od ostatních fragmentů (vnitřní tryptické peptidy). TAILS izoluje N-koncové peptidy identifikací a odstraněním vnitřních tryptických peptidů. Tato negativní selekce umožňuje metodě TAILS detekovat všechny N-konce v daných vzorcích. Alternativní metody, které se při identifikaci N-koncových peptidů spoléhají na volnou aminoskupinu N-konce, nemohou detekovat některé N-konce, protože jsou „přirozeně blokovány“ (tj. Přirozený protein nemá volnou aminoskupinu).
Metoda TAILS má řadu aplikací včetně identifikace nových substráty a proteázy (včetně těch, které mají neznámou a širokou specifičnost)[1] a jako způsob definování konců proteinů, který umožňuje anotaci proteinů. TAILS lze také použít k propojení proteáz s řadou definovaných biologických cest u nemocí, jako je rakovina za účelem jasnějšího pochopení substrátů a proteáz zapojených do chorobného stavu.[2]
Metoda
TAILS je 2D nebo 3D proteomický test pro značení a izolaci N-terminálu peptidy, vyvinutý skupinou na University of British Columbia.[1] Metoda TAILS je navržena pro srovnání více buněk ošetřených proteázou a kontrolních proteomových buněk.[2] Vzorky mohou být získány z různých zdrojů, včetně tkání, fibroblastů, rakovinných buněk a z tekutých výpotků.
Tento test izoluje N-koncové peptidy odstraněním vnitřních tryptických peptidů pomocí ultrafiltrace ponechání značených zralých N-koncových a neo-N-koncových peptidů k analýze tandemem hmotnostní spektrometrie (MS / MS). Tato negativní selekce umožňuje metodě TAILS detekovat všechny N-konce v daných vzorcích. Alternativní metody, které se při izolaci N-koncových peptidů spoléhají na volnou aminoskupinu N-konce, nemohou detekovat přirozeně blokované N-konce, protože nemají volnou aminoskupinu.
TAILS vyžaduje pro experimentování pouze malý vzorek peptidu (100–300 ug), lze jej použít s proteázami, které mají neznámou nebo širokou specificitu a podporují různé metody značení vzorků. Identifikuje však ~ 50% proteinů pomocí dvou nebo více různých a jedinečných peptidů (jeden z původního zralého N-konce a / nebo jeden nebo více neo-N-koncových peptidů prostřednictvím štěpného místa), které nepředstavují nezávislé biologické události, a proto nelze průměrovat pro kvantifikaci[je zapotřebí objasnění ]. Rovněž má potíže s validací výsledků pro analýzu N-terminomu na bázi jednoho peptidu[je zapotřebí objasnění ].[1]
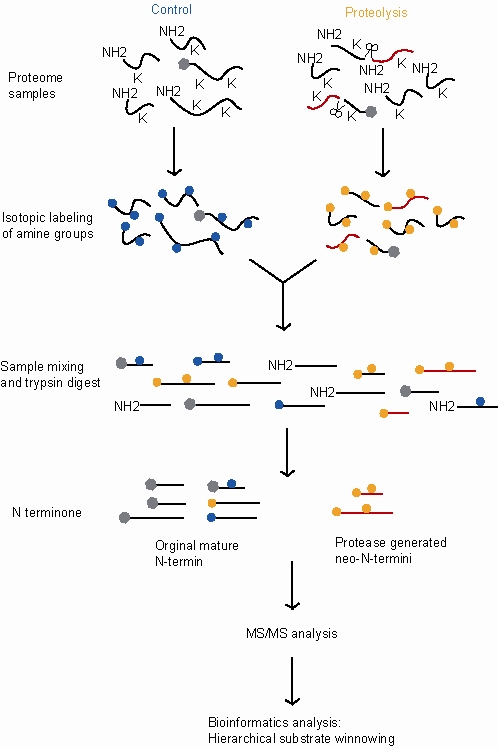
Následující kroky se týkají testu dimethylace-TAILS, porovnání kontrolního vzorku (vykazujícího normální proteolytickou aktivitu) a ošetřeného vzorku (který v tomto příkladu vykazuje další proteolytickou aktivitu).
- Celý protein proteolýza vyskytující se jak v ošetřených, tak v kontrolních vzorcích s další proteolytickou aktivitou v ošetřeném vzorku.
- Inaktivace proteáz a denaturace a redukce bílkovin.
- Označení stabilními izotopy. To umožňuje odlišit peptidy, které pocházejí z kontrolního vzorku, od peptidů, které pocházejí z ošetřeného vzorku, takže lze porovnat jejich relativní množství. V tomto příkladu se značení aplikuje redukční dimethylací primárních aminů za použití buď těžkého (d (2) C13) -formaldehydu pro ošetřené vzorky nebo lehkého (d (0) C12) -formaldehydu pro kontroly. Tato reakce je katalyzována kyanoborohydridem sodným a váže označené methylové skupiny na lysin-aminy a volné (∝) - aminoskupiny na N-koncích proteinů a produktů štěpení proteázou.
- Blokování reaktivních aminoskupin. To umožňuje identifikovat interní tryptické peptidy později v procesu, protože budou jedinými peptidy s reaktivními aminoskupinami. V tomto příkladu značkovací reakce (reduktivní dimethylace) také blokuje reaktivní aminoskupiny.
- Sdružování. Dva značené proteomy jsou nyní smíchány. Tím je zajištěno, že se vzorky ve všech následujících krocích zachází stejně, což umožňuje přesnější měření relativního množství proteinů ve dvou vzorcích.
- Trypsinizace. Tím se každý protein rozbije na fragmenty. Značené N-konce původních proteinů zůstávají blokovány, zatímco nové vnitřní tryptické peptidy mají volný N-konec.
- Negativní výběr. Do vzorku se přidá hyperrozvětvený polymer polyglycerol a aldehyd (HPG) specifický pro vazbu tryptických peptidů a reaguje s nově vytvořenými tryptickými peptidy prostřednictvím jejich volných N-konců. Stejně jako v kroku 3 výše je tato reakce katalyzována kyanoborohydridem sodným. Dimetylované lysinové acetylované a izotopově značené proteinové peptidy a neo (nové) -N-koncové peptidy jsou nereaktivní a zůstávají nevázané a lze je oddělit od komplexů poly-interních tryptických peptidů pomocí ultrafiltrace.
- The vymýt nenavázané proteiny jsou vysoce koncentrované s N-koncovými peptidy a neo-N-koncovými peptidy.
- Tento eluovaný vzorek je poté kvantifikován a analýza dokončena MS / MS.
- Poslední krok v TAILS zahrnuje bioinformatika. Použití hierarchického procesu převíjení substrátu, který odlišuje produkty proteolýzy pozadí a neštěpené proteiny kvantifikací izotopů peptidů a určitými kritérii bioinformatického vyhledávání.[1][2]
Typy
Typy OCASŮ se liší v metodách používaných k blokování a značení aminoskupin proteinů a produktů štěpení proteázou. Tyto aminoskupiny zahrnují lysin-aminy a volné (∝) -amino skupiny N-konců proteinů.
Dimethylační-TAILS postup je postup založený na chemickém značení, který se provádí v jednom kroku za použití amin-reaktivních izotopových reagencií. Označení dvou vzorků používá buď 12CH2-formaldehyd (světlo) nebo 13CD2-formaldehyd (těžký) a jako katalyzátor používá kyanoborohydrid sodný.[1] Výhodou této metody je, že je robustní, efektivní a nákladově efektivní. Postup značení u kontrol a vzorků ošetřených proteázou musí být proveden samostatně, než mohou být sloučeny, a je omezen na dva vzorky na experiment, což může být nevýhodou, pokud je třeba studovat více vzorků současně.[1]
Stabilní izotop značení aminokyselinami v buněčné kultuře (SILAC) je postup, který lze provést in vivo. Tento postup lze použít ve všech laboratořích buněčných kultur a jedná se o rutinně používanou techniku značení. Tento metabolické značení umožňuje inhibici dané proteázy v biologických vzorcích a analýzu ex vivo zpracovává se.[1] Výhodou použití této metody metabolického značení oproti chemickému značení je to, že umožňuje spolehlivou, rychlou a účinnou diskriminaci mezi proteiny odvozenými od skutečných buněk, které jsou vyšetřovány z kontaminantů, jako jsou proteiny v séru. SILAC TAILS lze použít pro analýzu až pěti multiplex Vzorky. SILAC není vhodný pro klinicky relevantní lidské vzorky, které nelze metabolicky označit. SILAC je drahá metoda a pro většinu laboratoří nemusí být proveditelnou volbou.[1]
The izobarická značka pro relativní a absolutní kvantifikaci Metoda (iTRAQ) nebo iTRAQ-TAILS umožňuje kvantifikaci více vzorků současně. Tato metoda má schopnost simultánně analyzovat od 4 do 8 vzorků v multiplexních experimentech s použitím reagencií iTRAQ se čtyřmi a osmi plexy. Tato metoda poskytuje vysokou přesnost identifikace a kvantifikace vzorků a umožňuje reprodukovatelnější analýzu replikátů vzorků.[1] Stejně jako ostatní metody iTRAQ vyžaduje iTRAQ-TAILS hmotnostní spektrometr MALDI a nákladná činidla iTRAQ.
Alternativní metody
Existuje několik alternativních přístupů ke studiu produktů N-konců a proteolýzy.
Acetylace aminů následovaná tryptickým štěpením a biotinylací volných N-koncových peptidů používá chemickou (acetylaci) k označení volných lysinů a N-konců. Blokované N-konce jsou poté negativně vybrány. Přirozeně volné vnitřní N-konce a blokované N-konce však nelze po acetylaci rozlišit. Tato metoda nepoužívá izotopové značení, takže je obtížné kvantifikovat zjištění. Také je těžké rozlišovat mezi experimentálními produkty a produkty proteolýzy pozadí.[3]
Guanidace lysinu následovaná biotinylací N-konců používá chemickou látku k blokování lysinových zbytků a značení volných N-konců. Poté jsou vybrány označené volné N-konce. Nevýhodou této metody je, že nálezy nelze aplikovat na statistický model využívající neštěpené peptidy kvůli tomu, že nejsou schopny zachytit přirozeně blokované N-konce. Protože nezahrnuje izotopové značení, nelze výsledky kvantifikovat. Je také známo, že místo štěpení lze označovat.[4]
Subtiligázová biotinylace N-konců využívá enzymatické značení N-koncových peptidů, ale nepoužívá chemikálie blokující lysin. Bez blokování lysinu bude mnoho štěpeného N-koncového peptidu příliš krátké na identifikaci. Výsledky mohou být vysoce závislé na vlastnostech subtiligázy, takže mohou být zkreslené. Tato metoda nezachycuje přirozeně blokované N-konce a také nepoužívá izotopové značení, takže by bylo obtížné kvantifikovat nálezy.[5]
Označení N-konců ITRAQ používá k označování N-konců iTRAQ. Peptidy Ne-N-konců jsou vybírány in silico. Nevýhodou této techniky je, že je zapotřebí hmotnostní spektrometr MALDI a požadovaná činidla iTRAQ jsou nákladná. Tato metoda nezachycuje přirozeně blokované N-konce. Celý proces bude vyžadovat 50-100 mg peptidových vzorků.[6]
Kombinovaná frakční diagonální chromatografie (COFRADIC) umožňuje různé značení pro přirozeně blokované N-konce a neo-N-konce generované proteázou. Všechny blokované N-konce jsou negativně vybrány. Proces však vyžaduje mnoho chemického zpracování, chromatografie a hmotnostní spektrometrie. Nejlepší výsledky separace závisí na modifikaci aminokyselin, jako je oxidace methioninu, k níž nedochází během manipulace. Tato metoda vyžaduje 150 MS / MS analýz na vzorek, ale vzorky lze shromáždit pro hmotnostní spektrometrii (a počet analýz lze snížit). Tato technika je vhodná pro použití pro proteázy s neznámou nebo širokou specificitou.[7]
Viz také
Reference
- ^ A b C d E F G h i Kleifeld, Oded; Doucet, Alain; Prudová, Anna; Auf Dem Keller, Ulrich; Gioia, Magda; Kizhakkedathu, Jayachandran N; Celkově Christopher M (2011). "Identifikace a kvantifikace proteolytických událostí a přirozeného N terminomu pomocí izotopového značení substrátů terminálním aminem". Přírodní protokoly. 6 (10): 1578–611. doi:10.1038 / nprot.2011.382. PMID 21959240.
- ^ A b C Kleifeld, Oded; Doucet, Alain; Auf Dem Keller, Ulrich; Prudová, Anna; Schilling, Oliver; Kainthan, Rajesh K; Starr, Amanda E; Foster, Leonard J; et al. (2010). „Izotopové značení terminálních aminů ve složitých vzorcích identifikuje produkty štěpení proteinovými N-konci a proteázou“. Přírodní biotechnologie. 28 (3): 281–8. doi:10.1038 / nbt.1611. PMID 20208520.
- ^ McDonald, Lucy; Robertson, Duncan HL; Hurst, Jane L; Beynon, Robert J (2005). „Poziční proteomika: Selektivní výtěžek a analýza N-koncových proteolytických peptidů“. Přírodní metody. 2 (12): 955–7. doi:10.1038 / nmeth811. PMID 16299481.
- ^ Timmer, John C .; Enoksson, Mari; Wildfang, Eric; Zhu, Wenhong; Igarashi, Yoshinobu; Denault, Jean-Benard; Ma, Yuliang; Dummitt, Benjamin; et al. (2007). "Profilování konstitutivních proteolytických událostí in vivo". Biochemical Journal. 407 (1): 41–8. doi:10.1042 / BJ20070775. PMC 2267409. PMID 17650073.
- ^ Mahrus, Sami; Trinidad, Jonathan C .; Barkan, David T .; Sali, Andrej; Burlingame, Alma L .; Wells, James A. (2008). „Globální sekvenování proteolytických štěpných míst v apoptóze specifickým označením proteinů N Termini“. Buňka. 134 (5): 866–76. doi:10.1016 / j.cell.2008.08.012. PMC 2566540. PMID 18722006.
- ^ Enoksson, Mari; Li, Jingwei; Ivancic, Melanie M .; Timmer, John C .; Wildfang, Eric; Eroshkin, Alexey; Salvesen, Guy S .; Tao, W. Andy (2007). "Identifikace proteinových štěpných míst kvantitativní proteomikou". Journal of Proteome Research. 6 (7): 2850–8. doi:10.1021 / pr0701052. PMID 17547438.
- ^ Gevaert, Kris; Goethals, Marc; Martens, Lennart; Van Damme, Jozef; Staes, An; Thomas, Grégoire R .; Vandekerckhove, Joël (2003). „Zkoumání proteomů a analýza zpracování proteinů hmotnostní spektrometrickou identifikací tříděných N-koncových peptidů“. Přírodní biotechnologie. 21 (5): 566–9. doi:10.1038 / nbt810. PMID 12665801.