CLE peptid - CLE peptide - Wikipedia
CLE peptidy (CLAVATA3 / Embryo související s okolním regionem) jsou skupina peptidy nalezené v rostlinách, které se na něm podílejí buněčná signalizace. Produkce je řízena geny CLE. Po navázání na CLE peptid receptor v jiné buňce dochází k řetězové reakci událostí, která může vést k různým fyziologickým a vývojovým procesům. Tato signální cesta je konzervována různorodě suchozemské rostliny.[1]
Pozadí
Rostliny i zvířata používají oba malé polypeptidy pro signalizaci v komunikaci mezi buňkami. CLAVATA3 / Embryo Surrounding Region-Related, také známý jako rostlinný peptidový hormon, je signalizace důležitá pro buněčnou signalizaci, ale také na dálkovou komunikaci. Tyto dvě akce jsou zvláště důležité pro rostlinné buňky, protože jsou stacionární a musí provádět expanzi buněk. V mnohobuněčných organismech bylo shledáno, že komunikace mezi buňkami je velmi důležitá pro mnoho růstových procesů, ke kterým dochází uvnitř organismu. Polypeptidy s 12 nebo 13 aminokyselinami jsou zralé formy proteinů CLE, které jsou odvozeny od konzervovaných domén CLE.[2][3][4] Stále více genů CLE je identifikováno a v této oblasti probíhá další výzkum. Geny CLE nebyly nalezeny pouze v semenných rostlinách, ale také v lykofyty, mechorosty, a zelené řasy.[5][6]
Geny
Většina výzkumů, které byly prováděny na signalizaci peptidů CLE, byla provedena s Arabidopsis, protože tento genom obsahuje 32 členů rodiny genů CLE. CLV3, který patří do rodiny genů CLE, se nachází v jedné nebo více tkáních Arabidopsis. Všech 32 členů rodiny CLE sdílí dvě charakteristiky, které zahrnují: kódování malého proteinu s domnělým sekrečním signálem na jejich N-koncích a obsahují konzervovaný motiv CLE na jejich C-koncích nebo v jejich blízkosti.[7] 32 členů rodiny genů CLE pocházelo z mutací původního genu.
Struktury
CLE peptidy jsou kódovány geny CLE. Tyto peptidy se liší strukturou, přičemž každá peptidová struktura provádí v rostlině jinou práci. Bylo zjištěno, že minimální délka fungujících peptidů CLE je 12 aminokyseliny s několika kritickými zbytky.[8] V rostlině se nacházejí dvě různé peptidové struktury, které jsou typu A a typu B. Když jsou hormony typu A vylučovány, rostlina zpomaluje rychlost růstu kořenů, zatímco sekrece peptidů typu B ovlivňuje cévní růst rostliny.[9] Sekrece peptidů typu A urychluje vaskulární vývoj rostliny, který je zprostředkován peptidy typu B. To naznačuje, že tyto dva typy peptidů spolupracují na regulaci růstu rostliny. Specifické peptidy jsou:[8]
Peptidy typu A.
- CLE 1/3/4
- CLE 2
- CLE 5/6
- CLE 7
- CLE 8
- CLE 9
- CLE 10
- CLE 11
- CLE 12
- CLE 13
- CLE 14
- CLE 16
- CLE 17
- CLE 18
- CLE 19
- CLE 20
- CLE 21
- CLE 22
- CLE 25
- CLE 26
- CLE 27
- CLE 40
- CLE 45
Peptidy typu B.
- CLE 41/44 / TDIF
- CLE 42
- CLE 43
- CLE 46
Signalizace v natáčení apikálního meristému

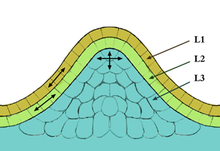
Meristematické buňky dávají vzniknout různým orgánům rostliny a udržují rostlinu. Existují dva typy meristematických tkání 1) Apikální meristém 2) Laterální meristém. Apikální meristém je dvou typů; výhonkový apikální meristém (SAM) dává vznik orgánům, jako jsou listy a květy, zatímco kořenový apikální meristém (RAM) poskytuje meristematické buňky pro budoucí růst kořenů. Buňky SAM a RAM se rychle dělí a jsou považovány za neurčité, protože nemají žádný definovaný konečný stav. V tomto smyslu jsou meristematické buňky často srovnávány s kmenovými buňkami u zvířat, která mají analogické chování a funkci. V rostlinách hrají buňky SAM hlavní roli v celkovém růstu a vývoji, je to způsobeno skutečností, že všechny buňky tvořící hlavní části rostliny pocházejí z výhonkového apikálního meristému (SAM). V rámci SAM se nacházejí tři různé důležité oblasti a zahrnují centrální zónu, periferní zónu) a meristém žeber. Každá z těchto oblastí hraje důležitou roli v produkci nových kmenových buněk v rámci SAM. Všechny SAM mají obvykle kopulovitý tvar a mají struktury, které jsou vrstvené a jsou popsány jako tunica a korpus. CLV3 hraje důležitou roli při regulaci produkce kmenových buněk v oblasti centrální zóny (SAM), to platí také pro buňku podporující gen WUSCHEL (WUS). Kombinace těchto dvou genů reguluje produkci kmenových buněk pomocí WUS negativně nebo pozitivně reguluje produkci kmenových buněk kontrolou genu CLV3 .;[10][11]
Geny v jiných rostlinách
Geny CLE byly nalezeny v řadě jednoděložní rostliny, dvouděložné rostliny, a dokonce mech. Výzkum dokonce ukázal, že některé rostliny jako rýže obsahují přítomnost multi-CLE domény.[5][7] V genomech genu byly také nalezeny různé geny podobné CLE háďátka parazitující na rostlinách jako řepa, sója a bramborové cystotvorné hlístice.[12][5][13]
Reference
- ^ Betsuyaku S, Sawa S, Yamada M (2011). „Funkce peptidů CLE při vývoji rostlin a interakcích rostlin a mikrobů“. Kniha Arabidopsis. 9: e0149. doi:10.1199 / tab.0149. PMC 3268505. PMID 22303273.
- ^ Kondo T, Sawa S, Kinoshita A, Mizuno S, Kakimoto T, Fukuda H, sakagami Y (2006). "Rostlinný peptid kódovaný pomocí CLV3 identifikovaný in situ MALDI-TOF MS analýzou". Věda. 313 (5788): 845–48. doi:10.1126 / science.1128439. JSTOR 3846935.
- ^ Ito Y, Nakanomyo I, Motose H, Iwamoto K, Sawa S, Dohmae N, Fukuda H (srpen 2006). "Dodeca-CLE peptidy jako supresory diferenciace rostlinných kmenových buněk". Věda. 313 (5788): 842–5. doi:10.1126 / science.1128436. PMID 16902140.
- ^ Ohyama K, Shinohara H, Ogawa-Ohnishi M, Matsubayashi Y (srpen 2009). "Glykopeptid regulující osud kmenových buněk v Arabidopsis thaliana". Nat. Chem. Biol. 5 (8): 578–80. doi:10.1038 / nchembio.182. PMID 19525968.
- ^ A b C Oelkers K, Goffard N, Weiller GF, Gresshoff PM, Mathesius U, Frickey T (leden 2008). „Bioinformatická analýza rodiny signálních peptidů CLE“. BMC Plant Biol. 8: 1. doi:10.1186/1471-2229-8-1. PMC 2254619. PMID 18171480.
- ^ Miwa H, Tamaki T, Fukuda H, Sawa S (červen 2009). „Vývoj signalizace CLE: počátky rozmanitosti receptorů CLV1 a SOL2 / CRN“. Plant Signal Behav. 4 (6): 477–81. doi:10,4161 / psb.4.6.8391. PMC 2688290. PMID 19816140.
- ^ A b Cock JM, McCormick S (červenec 2001). „Velká rodina genů, které sdílejí homologii s CLAVATA3“. Plant Physiol. 126 (3): 939–42. doi:10,1104 / pp.126.3.939. PMC 1540125. PMID 11457943.
- ^ A b Yamaguchi YL, Ishida T, Sawa S (2016). „CLE peptidy a jejich signální dráhy ve vývoji rostlin“. J. Exp. Bot. 67 (16): 4813–26. doi:10.1093 / jxb / erw208. PMID 27229733.
- ^ Whitford R, Fernandez A, De Groodt R, Ortega E, Hilson P (listopad 2008). "Rostlinné CLE peptidy ze dvou odlišných funkčních tříd synergicky indukují dělení vaskulárních buněk". Proc. Natl. Acad. Sci. USA. 105 (47): 18625–30. doi:10.1073 / pnas.0809395105. PMC 2587568. PMID 19011104.
- ^ Značka U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R (červenec 2000). "Závislost osudu kmenových buněk u Arabidopsis na zpětnovazební smyčce regulované aktivitou CLV3". Věda. 289 (5479): 617–9. doi:10.1126 / science.289.5479.617. PMID 10915624.
- ^ Schoof H, Lenhard M, Haecker A, Mayer KF, Jürgens G, Laux T (březen 2000). "Populace kmenových buněk Arabidopsis střílí meristémy udržované regulační smyčkou mezi geny CLAVATA a WUSCHEL". Buňka. 100 (6): 635–44. doi:10.1016 / s0092-8674 (00) 80700-x. PMID 10761929.
- ^ Wang X, Mitchum MG, Gao B, Li C, Diab H, Baum TJ, Hussey RS, Davis EL (březen 2005). „Parazitický gen z hlístice parazitické na rostlině s funkcí podobnou CLAVATA3 / ESR (CLE) z Arabidopsis thaliana“. Mol. Plant Pathol. 6 (2): 187–91. doi:10.1111 / j.1364-3703.2005.00270.x. PMID 20565649.
- ^ Guo Y, Han L, Hymes M, Denver R, Clark SE (září 2010). „CLAVATA2 tvoří odlišný komplex vazebných receptorů CLE regulující specifikaci kmenových buněk Arabidopsis“. Rostlina J. 63 (6): 889–900. doi:10.1111 / j.1365-313X.2010.04295.x. PMC 2974754. PMID 20626648.
Další čtení
- Bommert P, Je BI, Goldshmidt A, Jackson D (říjen 2013). "Kukuřičný gen Gα COMPACT PLANT2 funguje v signalizaci CLAVATA a reguluje velikost meristému výhonku". Příroda. 502 (7472): 555–8. doi:10.1038 / příroda1283. PMID 24025774.
- Chu H, Liang W, Li J, Hong F, Wu Y, Wang L, Wang J, Wu P, Liu C, Zhang Q, Xu J, Zhang D (prosinec 2013). „Signální modul CLE-WOX reguluje údržbu kořenového meristému a vývoj vaskulární tkáně v rýži“. J. Exp. Bot. 64 (17): 5359–69. doi:10.1093 / jxb / ert301. PMID 24043854.
- Clark SE, Williams RW, Meyerowitz EM (květen 1997). „CLAVATA1 gen kóduje domnělou receptorovou kinázu, která řídí velikost výhonku a květního meristému u Arabidopsis“. Buňka. 89 (4): 575–85. doi:10.1016 / s0092-8674 (00) 80239-1. PMID 9160749.
- Jun J, Fiume E, Roeder AH, Meng L, Sharma VK, Osmont KS, Baker C, Ha CM, Meyerowitz EM, Feldman LJ, Fletcher JC (prosinec 2010). „Komplexní analýza exprese a aktivity nadměrné exprese signálního genu polypeptidu CLE u Arabidopsis“. Plant Physiol. 154 (4): 1721–36. doi:10.1104 / pp.110.163683. PMC 2996011. PMID 20884811.
- Laux T, Mayer KF, Berger J, Jürgens G (leden 1996). „Gen WUSCHEL je vyžadován pro integritu výhonku a květního meristému u Arabidopsis.“ Rozvoj. 122 (1): 87–96. PMID 8565856.